Working Group 2 - Chapter 15: Polar Regions (Arctic and Antarctic) - (AR4-WG2-15)
Original at: http://www.ipcc.ch/publications_and_data/ar4/wg2/en/ch15.html
Main AR4 Index | Working Group WG2 Index | Table of Contents | Authors | Executive Summary | Annotated Text | References | Reviewer Comments
With the exception of Chapter and Section headings, all coloured text has been inserted by AccessIPCC. The non-coloured text is the IPCC original.
A number of emails from the Climate Research Unit (CRU) of the University of East Anglia were published on the Internet in November 2009. This has provided a window into the world of climate science.
We have identified a number of key individuals involved in the emails whom we have designated as Persons of Concern [PoC]; a Journal in which a PoC has published has been designated as a Journal of Concern [JoC].
This is not to suggest that we believe such papers are necessarily flawed, but rather that, as Joseph Alcamo noted at Bali in October 2009, "as policymakers and the public begin to grasp the multi-billion dollar price tag for mitigating and adapting to climate change, we should expect a sharper questioning of the science behind climate policy".
References occur in a list at the end of each chapter. Citations are within the normal text of sections and paragraphs.
| Tag | Explanation | Where Used | References | Citations |
|---|---|---|---|---|
| PoC |
Person of Concern Key individual involved in CRU emails as defined in this spreadsheet. |
References, Citations, IPCC Roles | 9 | 19 |
| JoC |
Journal of Concern A Journal which has published articles by one or more PoCs (Person of Concern) |
References, Citations | 105 | 134 |
| MoS |
Model or Simulation Reference appears to be a model or simulation, not observation or experiment |
References, Citations | 29 | 36 |
| NPR |
Non Peer Reviewed Reference has no Journal or no Volume or no Pages or it has Editors. |
References, Citations | 114 | 202 |
| SRC |
Self Reference Concern Author of a chapter containing references to own work. |
References, Citations, IPCC Roles | 74 | 163 |
| ARC |
Paper authored or co-authored by person who is also in list of Authors of another chapter. |
References, Citations | 24 | 45 |
| 2007 |
Paper dated 2007, when IPCC policy stated cutoff was December 2005 |
References, Citations | 11 | 20 |
| Ambiguous |
The short inline citation matched with more than one reference; however, AccessIPCC will link to the first reference found. |
Citations | - | 5 |
| NotFound |
The short inline citation was not matched with any reference. Believed to be caused by typing errors. |
Citations | - | 5 |
| Clean |
The reference was probably peer reviewed. |
References, Citations | 93 | 112 |
Coordinating Lead Authors:
Oleg A. Anisimov (Russia) [SRC:10], David G. Vaughan (UK) [SRC:6],
| Concern | Occurrence |
|---|---|
| SRC >= 5 | 2 |
| Potentially Biased Authors | 2 |
Lead Authors:
Terry Callaghan (Sweden/UK) [SRC:13], Christopher Furgal (Canada) [SRC:6], Harvey Marchant (Australia) [SRC:1], Terry D. Prowse (Canada) [SRC:17], Hjalmar Vilhjálmsson (Iceland) [SRC:2], John E. Walsh (USA) [SRC:7], Contributing Authors: Torben R. Christensen (Sweden), Donald L. Forbes (Canada) [SRC:1], Frederick E. Nelson (USA) [SRC:5], Mark Nuttall (Canada/UK) [SRC:3], James D. Reist (Canada) [SRC:9], George A. Rose (Canada) [SRC:3], Jef Vandenberghe (The Netherlands) [SRC:3], Fred J. Wrona (Canada) [SRC:10],
| Concern | Occurrence |
|---|---|
| SRC >= 5 | 7 |
| SRC 1-4 | 6 |
| Potentially Biased Authors | 13 |
| Impartial Authors | 1 |
Review Editors:
Roger Barry (USA) [SRC:1], Robert Jefferies (Canada) [SRC:1], John Stone (Canada),
| Concern | Occurrence |
|---|---|
| SRC 1-4 | 2 |
| Potentially Biased Authors | 2 |
| Impartial Authors | 1 |
This chapter should be cited as:
Anisimov, O.A., D.G. Vaughan, T.V. Callaghan, C. Furgal, H. Marchant, T.D. Prowse, H. Vilhjálmsson and J.E. Walsh, 2007: Polar regions (Arctic and Antarctic). Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, M.L. Parry, O.F. Canziani, J.P. Palutikof, P.J. van der Linden and C.E. Hanson, Eds., Cambridge University Press, Cambridge, 653-685.
Executive summary
In both polar regions, there is strong evidence of the ongoing impacts of climate change on terrestrial and freshwater species, communities and ecosystems (very high confidence). Recent studies project that such changes will continue (high confidence), with implications for biological resources and globally important feedbacks to climate (medium confidence).
Strong evidence exists of changes in species’ ranges and abundances and in the position of some tree lines in the Arctic (high confidence). An increase in greenness and biological productivity has occurred in parts of the Arctic (high confidence). Surface albedo is projected to decrease and the exchange of greenhouse gases between polar landscapes and the atmosphere will change (very high confidence). Although recent models predict that a small net accumulation of carbon will occur in Arctic tundra during the present century (low confidence), higher methane emissions responding to the thawing of permafrost and an overall increase in wetlands will enhance radiative forcing (medium confidence). [ 15.4.1 , 15.4.2 , 15.4.6 ].
In both polar regions, components of the terrestrial cryosphere and hydrology are increasingly being affected by climate change (very high confidence). These changes will have cascading effects on key regional bio-physical systems and cause global climatic feedbacks, and in the north will affect socio-economic systems (high confidence).
Freshwater and ice flows into polar oceans have a direct impact on sea level and (in conjunction with the melt of sea ice) are important in maintaining the thermohaline circulation. In the Arctic, there has been increased Eurasian river discharge to the Arctic Ocean, and continued declines in the ice volume of Arctic and sub-Arctic glaciers and the Greenland ice sheet (very high confidence). Some parts of the Antarctic ice sheet are also losing significant volume (very high confidence). Changes to cryospheric processes are also modifying seasonal runoff and routings (very high confidence). These combined effects will impact freshwater, riparian and near-shore marine systems (high confidence) around the Arctic, and on sub-Antarctic islands. In the Arctic, economic benefits, such as enhanced hydropower potential, may accrue, but some livelihoods are likely to be adversely affected (high confidence). Adaptation will be required to maintain freshwater transportation networks with the loss of ice cover (high confidence). [ 15.4.1 , 15.4.6 , 15.7.1 ].
Continued changes in sea-ice extent, warming and acidification of the polar oceans are likely to further impact the biomass and community composition of marine biota as well as Arctic human activities (high confidence).
Although earlier claims of a substantial mid-20th century reduction of Antarctic sea-ice extent are now questioned, a recently reported decline in krill abundance is due to regional reductions in Antarctic sea-ice extent; any further decline will adversely impact their predators and ecosystems (high confidence). Acidification of polar waters is predicted to have adverse effects on calcified organisms and consequential effects on species that rely upon them (high confidence). The impact of climate change on Arctic fisheries will be regionally specific; some beneficial and some detrimental. The reduction of Arctic sea ice has led to improved marine access, increased coastal wave action, changes in coastal ecology/biological production and adverse effects on ice-dependent marine wildlife, and continued loss of Arctic sea ice will have human costs and benefits (high confidence). [ 15.2.1 , 15.2.2 , 15.4.3 ].
Already Arctic human communities are adapting to climate change, but both external and internal stressors challenge their adaptive capabilities (high confidence). Benefits associated with climate change will be regionally specific and widely variable at different locations (medium confidence).
Impacts on food accessibility and availability, and personal safety are leading to changes in resource and wildlife management and in livelihoods of individuals (e.g., hunting, travelling) (high confidence). The resilience shown historically by Arctic indigenous peoples is now being severely tested (high confidence). Warming and thawing of permafrost will bring detrimental impacts on community infrastructure (very high confidence). Substantial investments will be needed to adapt or relocate physical structures and communities (high confidence). The benefits of a less severe climate are dependant on local conditions, but include reduced heating costs, increasing agricultural and forestry opportunities, more navigable northern sea routes, and marine access to resources (medium confidence). [ 15.4.5 , 15.5 , 15.7 ]
15.1 Introduction
The polar regions are increasingly recognised as being:
- geopolitically and economically important,
- extremely vulnerable to current and projected climate change,
- the regions with the greatest potential to affect global climate and thus human populations and biodiversity.
Sub-regions of the Arctic and Antarctic have shown the most rapid rates of warming in recent years. Substantial environmental impacts of climate change show profound regional differences both within and between the polar regions, and enormous complexity in their interactions. The impacts of this climate change in the polar regions over the next 100 years will exceed the impacts forecast for many other regions and will produce feedbacks that will have globally significant consequences. However, the complexity of response in biological and human systems, and the fact that these systems are subject to multiple stressors, means that future impacts remain very difficult to predict.
15.1.1 Summary of knowledge assessed in the TAR
This chapter updates and extends the discussion of polar regions in the IPCC Working Group II, Third Assessment Report (TAR, Anisimov et al., 2001 [Ambiguous] ). That report summarised the climatic changes that have been observed in the polar regions over the 20th century, the impacts those changes have had on the environment, and the likely impact of projected climate change in the future. The following summarises the key findings of that assessment to which ‘very-high confidence’ or ‘high confidence’ was attached (see Figure 15.1 for overview and place names used in this chapter).
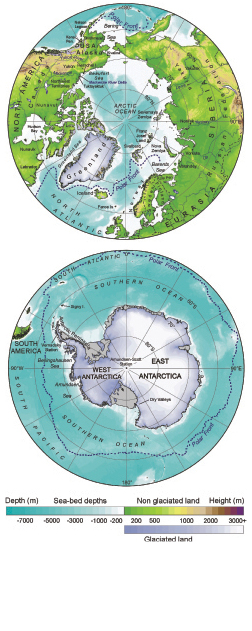
Figure 15.1. Location maps of the North and South polar regions, including place names used in the text. The topography of glaciated and non-glaciated terrain is shown by using different shading schemes. The polar fronts shown are intended to give an approximate location for the extent of cold, polar waters but are, in places, open to interpretation and fluctuations. (This and other maps were drawn by P. Fretwell, British Antarctic Survey.)
Key trends highlighted in the TAR
- In the Arctic, during the 20th century, air temperatures over extensive land areas increased by up to 5°C; sea ice thinned and declined in extent; Atlantic water flowing into the Arctic Ocean warmed; and terrestrial permafrost and Eurasian spring snow decreased in extent.
- There has been a marked warming in the Antarctic Peninsula over the last half-century. There has been no overall change in Antarctic sea-ice extent over the period 1973-1996.
Key regional projections highlighted in the TAR
- Increased melting of Arctic glaciers and the Greenland ice sheet, but thickening of the Antarctic ice sheet due to increased precipitation, were projected.
- Exposure of more bare ground and consequent changes in terrestrial biology on the Antarctic Peninsula were anticipated.
- Substantial loss of sea ice at both poles was projected.
- Reduction of permafrost area and extensive thickening of the active layer in the Arctic was expected to lead to altered landscapes and damage to infrastructure.
- Climate change combined with other stresses was projected to affect human Arctic communities, with particularly disruptive impacts on indigenous peoples following traditional and subsistence lifestyles.
- Economic costs and benefits were expected to vary among regions and communities.
Key polar drivers of global climate change identified in the TAR
- Changes in albedo due to reduced sea-ice and snow extent were expected to cause additional heating of the surface and further reductions in ice/snow cover.
Awareness of such issues led to the preparation of a uniquely detailed assessment of the impacts of climate change in the Arctic ( ACIA, 2005 [NPR] ), which has been drawn upon heavily in the Arctic component of this chapter. There is no similarly detailed report for the Antarctic.
15.2 Current sensitivity/vulnerability
15.2.1 Climate, environment and socio-economic state
Arctic
For several decades, surface air temperatures in the Arctic have warmed at approximately twice the global rate ( McBean et al., 2005 [NPR, ARC] ). The areally averaged warming north of 60°N has been 1-2°C since a temperature minimum in the 1960 s and 1970 s. In the marine Arctic, the 20th-century temperature record is marked by strong low-frequency (multi-decadal) variations ( Polyakov et al., 2002 [JoC] Serreze and Francis 2006 [JoC] ) have discussed the attribution of recent changes in terms of natural variability and anthropogenic forcing, concluding that a substantial proportion of the recent variability is circulation-driven, and that the Arctic is in the early stages of a manifestation of a human-induced greenhouse signature. This conclusion is based largely on the relatively slow rate of emergence of the greenhouse signal in model simulations of the late 20th and early 21st centuries.
The most recent ( 1980 to present) warming of much of the Arctic is strongest (about 1°C/decade) in winter and spring, and smallest in autumn; it is strongest over the interior portions of northern Asia and north-western North America ( McBean et al., 2005 [NPR, ARC] ). The latter regions, together with the Antarctic Peninsula, have been the most rapidly warming areas of the globe over the past several decades ( Turner et al., 2007 [JoC, SRC, 2007] ). The North Atlantic sub-polar seas show little warming during the same time period, probably because of their intimate connection with the cold, deep waters. Temperatures in the upper troposphere and stratosphere of the Arctic have cooled in recent decades, consistent with increases in greenhouse gases and with decreases in stratospheric ozone since 1979 ( Weatherhead et al., 2005 [NPR] ).
Precipitation in the Arctic shows signs of an increase over the past century, although the trends are small (about 1% per decade), highly variable in space, and highly uncertain because of deficiencies in the precipitation measurement network ( McBean et al., 2005 [NPR, ARC] ) and the difficulty in obtaining accurate measurements of rain and snow in windy polar regions. There is no evidence of systematic increases in intense storms in the Arctic ( Atkinson, 2005 [JoC, MoS] ) although coastal vulnerability to storms is increasing with the retreat of sea ice (see Section 15.4.6 ). Little is known about areally averaged precipitation over Greenland. The discharge of Eurasian rivers draining into the Arctic Ocean shows an increase since the 1930 s ( Peterson et al., 2002 [PoC, JoC, ARC] ), generally consistent with changes in temperature and the large-scale atmospheric circulation.
Reductions of Arctic sea ice and glaciers (see Lemke et al., 2007 [NPR, ARC, 2007] ), reductions in the duration of river and lake ice in much of the sub-Arctic ( Prowse et al., 2004 [NPR, SRC] ; Walsh et al., 2005 [NPR, PoC, SRC] ), and a recent ( 1980 s to present) warming of permafrost in nearly all areas for which measurements are available ( (Romanovsky et al., 2002; ) Walsh et al., 2005 [NPR, PoC, SRC] ) are consistent with the recent changes in Arctic surface air temperatures. Although there is visual evidence of permafrost degradation ( Lemke et al., 2007 [NPR, ARC, 2007] ), long-term measurements showing widespread thickening of the active layer are lacking. Changes in vegetation, particularly a transition from grasses to shrubs, has been reported in the North American Arctic ( Sturm et al., 2001 [JoC] ) and elsewhere ( Tape et al., 2006 [JoC] ), and satellite imagery has indicated an increase in the Normalised Difference Vegetation Index (NDVI, a measure of photosynthetically active biomass) over much of the Arctic ( Slayback et al., 2003 [JoC] ). This is consistent with a longer growing season and with documented changes in the seasonal variation in atmospheric CO2 concentrations as reported in the TAR. Broader ecosystem impacts of climate change in both polar regions are summarised by Walther et al. 2002 [JoC, ARC] ) and were documented more extensively for the Arctic by the Arctic Climate Impact Assessment ( ACIA, 2005 [NPR] ).
Recent analysis of air-borne data ( Krabill et al., 2004 [JoC] ), satellite data ( Howat et al., 2005 [JoC] ; Luckman et al., 2006 [JoC] ; Rignot and Kanagaratnam, 2006 [JoC] ) and seismic data ( Ekstrom et al., 2006 [JoC] ) indicate thinning around the periphery of the Greenland ice sheet, where summer melt has increased during the past 20 years ( Abdalati and Steffen, 2001 [JoC] ; Walsh et al., 2005 [NPR, PoC, SRC] ), while there is evidence of slower rates of thickening further inland ( Johannessen et al., 2005 [JoC] ).
The Arctic is now home to approximately 4 million residents ( Bogoyavlenskiy and Siggner, 2004 [NPR] ). Migration into the Arctic during the 20th century has resulted in a change of demographics such that indigenous peoples now represent 10% of the entire population. This influx has brought various forms of social, cultural and economic change ( Huntington, 1992 [NPR] ; Nuttall, 2000b [NPR, SRC] ). For most Arctic countries, only a small proportion of their total population lives in the Arctic, and settlement remains generally sparse ( Bogoyavlenskiy and Siggner, 2004 [NPR] ) and nomadic peoples are still significant in some countries. On average, however, two-thirds of the Arctic population live in settlements with more than 5,000 inhabitants. Indigenous residents have, in most regions, been encouraged to become permanent residents in fixed locations, which has had a predominantly negative effect on subsistence activities and some aspects of community health. At the same time, Arctic residents have experienced an increase in access to treated water supplies, sewage disposal, health care facilities and services, and improved transportation infrastructure which has increased access to such things as outside market food items ( Hild and Stordhal, 2004 [NPR] ). In general, the Arctic has a young, rapidly growing population with higher birth rates than their national averages, and rising but lower than national average life-expectancy. This is particularly true for indigenous populations, although some exceptions exist, such as in the Russian north, where population and life-expectancy has decreased since 1990 (Einarsson et al., 2004 [NotFound] ).
Political and administrative regimes in Arctic regions vary between countries. In particular, indigenous groups have different levels of self-determination and autonomy. Some regions (e.g., northern Canada and Greenland) now have formalised land-claim settlements, while in Eurasia indigenous claims have only recently begun to be addressed ( Freeman, 2000 [NPR] ). Wildlife management regimes and indigenous/non-indigenous roles in resource management also vary between regions. Nowadays, large-scale resource extraction initiatives and/or forms of social support play significant roles in the economies of many communities. Despite these changes, aspects of subsistence and pastoral livelihoods remain important.
Regardless of its small and dispersed population, the Arctic has become increasingly important in global politics and economies. For example, the deleterious effect on the health of Arctic residents of contaminants produced in other parts of the world has led to international agreements such as the Stockholm Convention on Persistent Organic Pollutants (Downey and Fenge, 2003 [NotFound] ). Furthermore, significant oil, gas and mineral resources (e.g., diamonds) are still to be developed in circum-Arctic regions that will further increase the importance of this region in the world (e.g., U.S. Geological Survey World Energy Assessment Team, 2000; Laherre, 2001 [NPR] ).
Antarctic
Direct measurements reveal considerable spatial variability in temperature trends in Antarctica. All meteorological stations on the Antarctic Peninsula show strong and significant warming over the last 50 years (see Section 15.6.3 ). However, of the other long-term (>30 years) mean annual temperature records available, twelve show warming, while seven show cooling; although only two of these (one of each) are significant at the 10% level ( Turner et al., 2005 [JoC] ). If the individual station records are considered as independent measurements, then the mean trend is warming at a rate comparable to mean global warming ( Vaughan et al., 2003 [JoC, SRC] ), but there is no evidence of a continent-wide ‘polar amplification’ in Antarctica. In some areas where cooling has occurred, such as the area around Amundsen-Scott Station at the South Pole, there is no evidence of directly attributable impacts, but elsewhere cooling has caused clear local impacts. For example, in the Dry Valleys, a 6 to 9% reduction in primary production in lakes and a >10%/yr decline in soil invertebrates has been observed ( Doran et al., 2002 [JoC, SRC] ). Although the impacts are less certain, precipitation has also declined on sub-Antarctic islands ( (Bergstrom and Chown, 1999 ) ).
Recent changes in Antarctic sea-ice extent are discussed in detail elsewhere ( Lemke et al., 2007 [NPR, ARC, 2007] ), but evidence highlighted in the TAR ( Anisimov et al., 2001 [Ambiguous] ) gleaned from records of whaling activities ( de la Mare, 1997 [JoC] ) is no longer considered reliable ( (Ackley et al., 2003 ) ). So, for the period before satellite observation, only direct local observations (e.g., ( Murphy et al., 1995 ) ) and proxies (e.g., Curran et al., 2003 [JoC] ) are available. For the satellite period ( 1978 to present) there has been no ubiquitous trend in Antarctic sea-ice duration, but there have been strong regional trends (see Figure 15.2 ). Sea-ice duration in the Ross Sea has increased, while in the Bellingshausen and Amundsen Seas it has decreased, with high statistical significance in each case ( (Parkinson, 2002; ) ( Zwally et al., 2002 ) ). This pattern strongly reflects trends in atmospheric temperature at nearby climate stations ( Vaughan et al., 2003 [JoC, SRC] ).
Increasing atmospheric CO2 concentrations are leading to an increased draw-down of CO2 by the oceans and, as a consequence, sea water is becoming more acidic (Royal Society, 2005 [NPR] ). As is the case in other parts of the world’s oceans, coccolithophorids and foraminifera are significant components of the pelagic microbial community of the Southern Ocean and contribute to the draw-down of atmospheric CO2 to the deep ocean. Experimental studies ( Riebesell et al., 2000 [JoC] ) indicate that elevated CO2 concentration reduces draw-down of CO2 compared with the production of organic matter. Recent investigations suggest that at the present rate of acidification of the Southern Ocean, pteropods (marine pelagic molluscs) will not be able to survive after 2100 and their loss will have significant consequences for the marine food web ( Orr et al., 2005 [PoC, JoC] ). Similarly, cold-water corals are threatened by increasing acidification. Furthermore, increasing acidification leads to changes in the chemistry of the oceans, altering the availability of nutrients and reducing the ability of the oceans to absorb CO2 from the atmosphere (Royal Society, 2005 [NPR] ; see also Chapter 4 this volume).
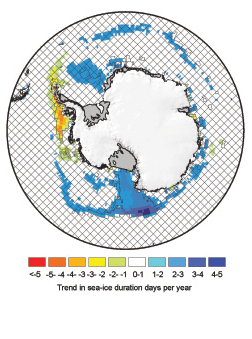
Figure 15.2. Trends in annual Antarctic sea-ice duration in days per year, for the period 1978 - 2004, after ( Parkinson 2002 ) ). Hatched areas show where trends are not significant at the 95% level. (Data compiled by W.M. Connolley, British Antarctic Survey.)
15.2.2 Vulnerability and adaptive capacity
Vulnerability is the degree to which a system is susceptible to, or unable to cope with, adverse effects of stress; whereas adaptive capacity, or resilience, is an ability to adjust to stress, to realise opportunities or to cope with consequences ( McCarthy et al., 2005 [NPR] ).
15.2.2.1 Terrestrial and marine ecosystems
Many polar species are particularly vulnerable to climate change because they are specialised and have adapted to harsh conditions in ways that are likely to make them poor competitors with potential immigrants from environmentally more benign regions (e.g., Callaghan et al., 2005 [NPR, SRC] ; Peck et al., 2006 [MoS] ). Other species require specific conditions, for example winter snow cover or a particular timing of food availability ( Mehlum, 1999 [NPR] ; Peck et al., 2006 [MoS] ). In addition, many species face multiple, concurrent human-induced stresses (including increased ultraviolet-B radiation, increasing contaminant loads, habitat loss and fragmentation) that will add to the impacts of climate change ( Walther et al., 2002 [JoC, ARC] ; McCarthy et al., 2005 [NPR] ).
Plants and animals in the polar regions are vulnerable to attacks from pests ( Juday et al., 2005 [NPR] ) and parasites ( (Albon et al., 2002; ) ( Kutz et al., 2002 ) ) that develop faster and are more prolific in warmer and moister conditions. Many terrestrial polar ecosystems are vulnerable because species richness is low in general, and redundancy within particular levels of food chains and some species groups is particularly low ( Matveyeva and Chernov, 2000 [NPR] ). Loss of a keystone species (e.g., lemmings, ( Turchin and Batzli, 2001 ) ) could have cascading effects on entire ecosystems.
Arctic
In Arctic ecosystems, adaptive capacity varies across species groups from plants that reproduce by cloning, which have relatively low adaptive potential, through some insects (e.g., ( Strathdee et al., 1993 ) ) that can adapt their life cycles, to micro-organisms that have great adaptive potential because of rapid turnover and universal dispersal. The adaptive capacity of current Arctic ecosystems is small because their extent is likely to be reduced substantially by compression between the general northwards expansion of forest, the current coastline and longer-term flooding of northern coastal wetlands as the sea level rises, and also as habitat is lost to land use (see Figure 15.3 ). General vulnerability to warming and lack of adaptive capacity of Arctic species and ecosystems are likely, as in the past, to lead to relocation rather than rapid adaptation to new climates (see Figure 15.3 ).
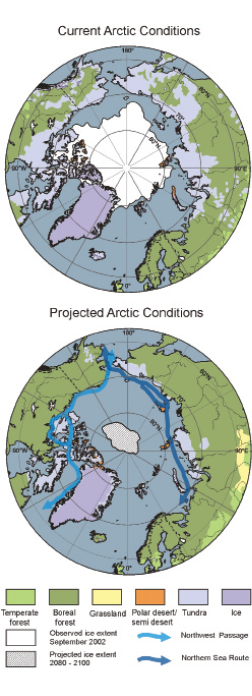
Figure 15.3. Present and projected vegetation and minimum sea-ice extent for Arctic and neighbouring regions. Vegetation maps based on floristic surveys (top) and projected vegetation for 2090 - 2100, predicted by the LPJ Dynamic Vegetation Model driven by the HadCM2 climate model (bottom) modified from Kaplan et al. 2003 [JoC, MoS, ARC] ) in Callaghan et al. 2005 [NPR, SRC] ). The original vegetation classes have been condensed as follows: grassland = temperate grassland and xerophytic scrubland; temperate forest = cool mixed forest, cool-temperate evergreen needle-leaved and mixed forest, temperate evergreen needle-leaved forest, temperate deciduous broadleaved forest; boreal forest = cool evergreen needle-leaved forest, cold deciduous forest, cold evergreen needle-leaved forest; tundra = low- and high-shrub tundra, erect dwarf-shrub tundra, prostrate dwarf-shrub tundra; polar desert/semi-desert = cushion forb, lichen and moss tundra. Also shown are observed minimum sea-ice extent for September 2002, and projected sea-ice minimum extent, together with potential new/improved sea routes (redrawn from Instanes et al., 2005 [NPR, SRC] ; Walsh et al., 2005 [NPR, PoC, SRC] ).
As air and sea water temperatures have increased in the Bering Sea, there have been associated changes in sea-ice cover, water-column properties and processes including primary production and sedimentation, and coupling with the bottom layer ( Grebmeier et al., 2006 [JoC] ). A change from Arctic to sub-Arctic conditions is happening with a northward movement of the pelagic-dominated marine ecosystem that was previously confined to the south-eastern Bering Sea. Thus communities that consist of organisms such as bottom-feeding birds and marine mammals are being replaced by communities dominated by pelagic fish. Changes in sea ice conditions have also affected subsistence and commercial harvests ( Grebmeier et al., 2006 [JoC] ).
Many Arctic and sub-Arctic seas (e.g., parts of the Bering and Barents Seas) are among the most productive in the world ( Sakshaug, 2003 [NPR] ), and yield about 7 Mt of fish per year, provide about US$15 billion in earnings ( Vilhjálmsson et al., 2005 [NPR, SRC] ), and employ 0.6 to 1 million people ( Agnarsson and Arnason, 2003 [NPR] ). In addition, Arctic marine ecosystems are important to indigenous peoples and rural communities following traditional and subsistence lifestyles ( Vilhjálmsson et al., 2005 [NPR, SRC] ).
Recent studies reveal that sea surface warming in the north-east Atlantic is accompanied by increasing abundance of the largest phytoplankton in cooler regions and their decreasing abundance in warmer regions ( Richardson and Schoeman, 2004 [JoC] ). In addition, the seasonal cycles of activities of marine micro-organisms and invertebrates and differences in the way components of pelagic communities respond to change, are leading to the activities of prey species and their predators becoming out of step. Continued warming is therefore likely to impact on the community composition and the numbers of primary and secondary producers, with consequential stresses on higher trophic levels. This will impact economically important species, primarily fish, and dependent predators such as marine mammals and sea birds ( Edwards and Richardson, 2004 [JoC, ARC] ).
Antarctic
Substantial evidence indicates major regional changes in Antarctic terrestrial and marine ecosystems in areas that have experienced warming. Increasing abundance of shallow-water sponges and their predators, declining abundances of krill, Adelie and Emperor penguins, and Weddell seals have all been recorded ( (Ainley et al., 2005 ) ). Only two species of native flowering plant, the Antarctic pearlwort (Colobanthus quitensis) and the Antarctic hair grass (Deschampsia antarctica) currently occur in small and isolated ice-free habitats on the Antarctic continent. Their increased abundance and distribution was ascribed to the increasing summer temperatures ( (Fowbert and Smith, 1994 ) ). Elsewhere on continental Antarctica, climate change is also affecting the vegetation, which is largely composed of algae, lichens and mosses, and changes are expected in future, as temperature, and water and nutrient availability, change ( Robinson et al., 2003 [JoC] ).
The marked reduction reported in the biomass of Antarctic krill (Euphausia superba) and an increase in the abundance of salps (principally Salpa thompsoni), a pelagic tunicate, may be related to regional changes in sea ice conditions ( Atkinson et al., 2004 [JoC] ). This change may also underlie the late-20th century changes in the demography of krill predators (marine mammals and sea birds) reported from the south-west Atlantic ( Fraser and Hoffmann, 2003 [MoS] ), and this connection indicates a potential vulnerability to climate change whose importance cannot yet be determined.
Recent studies on sub-Antarctic islands have shown increases in the abundance of alien species and negative impacts on the local biota such as a decline in the number and size of Sphagnum moss beds ( (Whinam and Copson, 2006 ) ). On these islands, increasing human activities and increasing temperatures are combining to promote successful invasions of non-indigenous species ( (Bergstrom and Chown, 1999 ) ).
15.2.2.2 Freshwater systems
Arctic
Climate variability/change has historically had, and will continue to have, impacts on Arctic freshwater resources. First-order impacts (e.g., changes to the snow/ice/water budget) play a significant role in important global climate processes, through feedbacks (e.g., changes to radiative feedbacks, stability of the oceanic stratification and thermohaline circulation, and carbon/methane source-sink status). Cascading effects have important consequences for the vulnerability of freshwater systems, as measured by their ecological or human resource value.
From an ecological perspective, the degree of vulnerability to many higher-order impacts (e.g., changes in aquatic geochemistry, habitat availability/quality, biodiversity) are related to gradual and/or abrupt threshold transitions such as those associated with water-phase changes (e.g., loss of freshwater ice cover) or coupled bio-chemical responses (e.g., precipitous declines in dissolved oxygen related to lake productivity) ( Wrona et al., 2005 [NPR, SRC] ). Historically, Arctic freshwater ecosystems have adapted to large variations in climate over long transitional periods (e.g., Ruhland and Smol, 2002 [MoS] ;( Ruhland et al., 2003 ) ), but in the next 100 years the combination of high-magnitude events and rapid rates of change will probably exceed the ability of the biota and their associated ecosystems to adapt ( Wrona et al., 2006a [SRC] ). This will result in significant changes and both positive and negative impacts. It is projected, however, that overall the negative effects will very probably outweigh the positive, implying that freshwater systems are vulnerable to climate change ( Wrona et al., 2005 [NPR, SRC] ).
From a human-use perspective, potential adaptation measures are extremely diverse, ranging from measures to facilitate modified use of the resource (e.g., changes in ice-road construction practices, increased open-water transportation, flow regulation for hydroelectric production, harvesting strategies, and methods of drinking-water access), to adaptation strategies to deal with increased/decreased freshwater hazards (e.g., protective structures to reduce flood risks or increase floods for aquatic systems ( Prowse and Beltaos, 2002 [SRC] ); changes to more land-based travel to avoid increasingly hazardous ice). Difficulties in pursuing adaptation strategies may be greatest for those who place strong cultural and social importance on traditional uses of freshwater resources ( McBean et al., 2005 [NPR, ARC] ; Nuttall et al., 2005 [NPR, SRC] ).
Antarctic
Antarctic freshwater systems are fewer and smaller than those in the Arctic, but are no less vulnerable to climate change. The microbial communities inhabiting these systems are likely to be modified by changing nutrient regimes, contaminants and introductions of species better able to cope with the changing conditions. A drop in air temperature of 0.7°C per decade late in the 20th century in the Dry Valleys led to a 6 to 9% drop in primary production in the lakes of the area ( Doran et al., 2002 [JoC, SRC] ). In marked contrast, summer air temperature on the maritime sub-Antarctic Signy Island increased by 1°C over the last 50 years and, over the period 1980 to 1995, water temperature in the lakes rose several times faster than the air temperature; this is one of the fastest responses to regional climate change in the Southern Hemisphere yet documented ( Quayle et al., 2002 [JoC] ). As a consequence, the annual ice-free period has lengthened by up to 4 weeks. In addition, the area of perennial snow cover on Signy Island has decreased by about 45% since 1951, and the associated change in microbial and geochemical processes has led to increased amounts of organic and inorganic nutrients entering the lakes. There has also been an explosion in the population of fur seals (Arctocephalus gazella) on the island due to decreased ice cover and increased area available for resting and moulting. Together these changes are leading to disruption of the ecosystem due to increased concentrations of nutrients (eutrophication) ( Quayle et al., 2003 [NPR] ). Similar ecological vulnerabilities are expected to exist in other Antarctic freshwater systems.
15.2.2.3 Permafrost
Permafrost, defined as sub-surface earth materials that remain at or below 0°C continuously for two or more years, is widespread in Arctic, sub-Arctic and high-mountain regions, and in the small areas of Antarctica without permanent ice cover. The physical processes of climate-permafrost interactions and observations of permafrost change are discussed elsewhere ( Lemke et al., 2007 [NPR, ARC, 2007] ); here we focus on the observed and projected changes of permafrost, and impacts they may have on natural and human systems in the Arctic.
Observational data are limited, but precise measurements in boreholes indicate that permafrost temperatures in the Arctic rose markedly during the last 50 years ( (Romanovsky et al., 2002 ) ), with rapid warming in Alaska ( Hinzman et al., 2005 [JoC, SRC] ), Canada ( (Beilman et al., 2001 ) ), Europe ( Harris et al., 2003 [ARC] ) and Siberia ( (Pavlov and Moskalenko, 2002 ) ). Short-term and localised warming associated with the reduction of snow cover ( Stieglitz et al., 2003 [JoC] ) and feedbacks associated with increased vegetation productivity ( Sturm et al., 2001 [JoC] ; Anisimov and Belolutskaia, 2004 [MoS, SRC] ; Chapin et al., 2005b [JoC] ) are, however, important considerations that must be taken into account.
In the context of the future climate change, there are two key concerns associated with the thawing of permafrost: the detrimental impact on the infrastructure built upon it, and the feedback to the global climate system through potential emission of greenhouse gases. These are discussed in Sections 15.7.1 and 15.4.2.3 .
15.2.2.4 Human populations
Neither Antarctica nor the sub-Antarctic islands have permanent human populations; the vast majority of residents are staff at scientific stations and summer-only visitors. While there are some areas of particular sensitivity (see Section 15.6.3 ), where climate change might dictate that facilities be abandoned, from a global perspective these can be viewed as logistical issues only for the organisations concerned.
In contrast, the archaeological record shows that humans have lived in the Arctic for thousands of years ( Pavlov et al., 2001 [JoC] ).
Previously, many Arctic peoples practised seasonal movements between settlements, and/or seasonally between activities (e.g., farming to fishing), and the semi-nomadic and nomadic following of game animals and herding. Today, most Arctic residents live in permanent communities, many of which exist in low-lying coastal areas. Despite the socio-economic changes taking place, many Arctic communities retain a strong relationship with the land and sea, with community economies that are a combination of subsistence and cash economies, in some cases, strongly associated with mineral, hydrocarbon and resource development ( Duhaime, 2004 [NPR] ). The vulnerable nature of Arctic communities, and particularly coastal indigenous communities, to climate change arises from their close relationship with the land, geographical location, reliance on the local environment for aspects of everyday life such as diet and economy, and the current state of social, cultural, economic and political change taking place in these regions.
Communities are already adapting to local environmental changes ( Krupnik and Jolly, 2002 [NPR] ; Nickels et al., 2002 [NPR, SRC] ) through wildlife management regimes, and changes in individual behaviours (i.e., shifts in timing and locations of particular activities) and they retain a substantial capacity to adapt. This is related to flexibility in economic organisation, detailed local knowledge and skills, and the sharing mechanisms and social networks which provide support in times of need ( (Berkes and Jolly, 2001 ) ). However, for some Arctic peoples, movement into permanent communities, along with shifts in lifestyle and culture, limits some aspects of adaptive capacity as more sedentary lifestyles minimise mobility, and increased participation in wage-economy jobs decreases the number of individuals able to provide foods from the local environment. The sustainability of this trend is unknown.
Small Arctic communities, however remote, are tightly tied politically, economically and socially to the national mainstream, as well as being linked to and affected by the global economy ( Nuttall et al., 2005 [NPR, SRC] ). Today, trade barriers, resource management regimes, political, legal and conservation interests, and globalisation all affect, constrain or reduce the abilities of Arctic communities to adapt to climate change ( Nuttall et al., 2005 [NPR, SRC] ). Trends in modernity within communities also affect adaptive capacity in both positive and negative ways. Increased access to outside markets and new technologies improve the ability to develop resources and a local economic base; however, increased time spent in wage-earning employment, while providing significant benefits at the individual and household levels through enhanced economic capacity, reduces time on the land observing and developing the knowledge that strengthens the ability to adapt. This underscores the reality that climate change is one of several interrelated problems affecting Arctic communities and livelihoods today ( Chapin et al., 2005a [NPR, SRC] ).
In some cases, indigenous peoples may consider adaptation strategies to be unacceptable, as they impact critical aspects of traditions and cultures. For example, the Inuit Circumpolar Conference has framed the issue of climate change in a submission to the United States Senate as an infringement on human rights because it restricts access to basic human needs as seen by the Inuit and will lead to the loss of culture and identity ( Watt-Cloutier, 2004 [NPR, MoS] ). Currently we do not know the limits of adaptive capacity among Arctic populations, or what the impacts of some adaptive measures will be.
15.3 Assumptions about future trends
15.3.1 Key regional impacts with importance to the global system
We expect climate change in the polar regions to have many direct regional impacts; however, those regional direct impacts may have global implications through the following processes and feedbacks.
- Reflectivity of snow, ice and vegetation: snow, ice and vegetation play vital roles in the global climate system, through albedo and insulation effects. Since the TAR, increasing evidence has emerged indicating a more rapid disappearance of snow and sea-ice cover in some areas (e.g., Siberia, Alaska, the Greenland Sea), and consequent changes of albedo may be leading to further climate change (e.g., Holland and Bitz, 2003).
- Retreat of glaciers and ice sheets, freshwater runoff, sea level and ocean circulation: the retreat of glaciers in the Arctic and more rapid melting of the edges of the Greenland ice sheet ( Section 15.2.1 ), together with observed increases in river runoff (Peterson et al., 2002), the major contributor, will alter the freshwater budget of the Arctic Ocean. Further changes are expected and could influence ocean circulation with global impacts (Lemke et al., 2007).
- Arctic terrestrial carbon flux: although models project that Arctic terrestrial ecosystems and the active layer will be a small sink for carbon in the next century, processes are complex and uncertainty is high. It is possible that increased emissions of carbon from thawing permafrost will lead to positive climate forcing (Sitch et al., 2007). Whether such emissions reach the atmosphere as methane or as carbon dioxide is important, because, on a per molecule basis, methane has more than 20 times the warming influence (Anisimov et al., 2005b).
- Migrating species: species that seasonally migrate from lower latitudes to polar regions rely on the existence of specific polar habitats, and if those habitats are compromised the effects will be felt in communities and food webs far beyond the polar regions. These habitats are likely to be compromised by direct or consequential climate change impacts such as drying of ponds and wetlands, and also by multiple stresses (e.g., land-use changes, hunting regulations).
- Methane hydrates: significant amounts of methane hydrates are contained in sediments, especially on Arctic continental shelves. As these areas warm, this methane may be released, adding to the greenhouse gas concentration in the atmosphere (Sloan, 2003; Maslin, 2004).
- Southern Ocean carbon flux: climate models indicate that stratification of the Southern Ocean will change. This could change the community structure of primary producers and alter rates of draw-down of atmospheric CO2 and its transport to the deep ocean.
15.3.2 Projected atmospheric changes
The areally averaged warming in the Arctic is projected to range from about 2°C to about 9°C by the year 2100, depending on the model and forcing scenario. The projected warming is largest in the northern autumn and winter, and is largest over the polar oceans in areas of sea-ice loss. Over land, the projected warming shows less seasonal variation, although regions such as the Canadian Archipelago are not well resolved.
In contrast to the unanimity of the models in predicting a north-polar amplification of warming, there are differences among the model projections concerning polar amplification in Antarctica, especially over the continent ( Parkinson, 2004 [MoS] ). However, in several simulations, the warming is amplified over a narrow Southern Ocean band from which sea ice retreats.
Global precipitation is projected to increase during the 21st century by 10 to 20% in response to the SRES [1] A1B emissions scenario of the IPCC Fourth Assessment Report (AR4) simulations. However, the seasonality and spatial patterns of the precipitation increase in the Arctic vary among the models. Similar results have emerged from other IPCC AR4 simulations ( Kattsov et al., 2007 [JoC, MoS, SRC, 2007] ). In addition, the partitioning among snow and rain will change in a warmer climate, affecting surface hydrology, terrestrial ecosystems and snow loads on structures. The ratio of rain to snow should increase especially in those seasons and Arctic sub-regions in which present-day air temperatures are close to freezing. The difference between precipitation and evapotranspiration (P-E), which over multi-year timescales is approximately equivalent to runoff (river discharge), is also projected to increase over the course of the 21st century. The projected increases of runoff by 2080 are generally in the range of 10 to 30%, largest in the A2 scenario and smallest in the B1 scenario. Of the major river basins, the largest increases are projected for the Lena River basin. Additional information on projected changes is presented elsewhere ( Meehl et al., 2007 [NPR, ARC, 2007] ). The effects of these freshwater changes on the thermohaline circulation are uncertain.
15.3.3 Projected changes in the oceans
A new study ( Zhang and Walsh, 2006 [JoC, MoS, SRC] ) based on the IPCC AR4 model simulations, projected mean reductions of annually averaged sea ice area in the Arctic by 2080 - 2100 of 31%, 33% and 22% under the A2, A1B and B1 scenarios, respectively (see Figure 15.3 ). A consistent model result is that the sea-ice loss is greater in summer than in winter, so that the area of seasonal (winter-only) sea-ice coverage actually increases in many models. The loss of summer sea ice will change the moisture supply to northern coastal regions and will be likely to impact the calving rates of glaciers that are now surrounded by sea ice for much of the year. There will also be increases in wind-driven transport and mixing of ocean waters in regions of sea-ice loss.
The projected increases of Arctic river discharge and precipitation over polar oceans, as well as the projections of increasing discharge from the Greenland ice sheet ( Lemke et al., 2007 [NPR, ARC, 2007] ), point to a freshening of the ocean surface in northern high latitudes. However, the projected changes of ice discharge (calving rates) are generally not available from the model simulations, since the ice sheet discharge is not explicitly included in coupled global models.
15.3.4 Projected changes on land
Arctic
Although seasonal snow cover on land is highly variable, it has important effects on the substrate and on local climate, primarily through its insulating properties and high albedo. In Eurasia, and to a lesser extent North America, there has been a persistent 5-6 day/decade increase in the duration of snow-free conditions over the past three decades ( (Dye, 2002 ) ). The reduction of snow residence time occurs primarily in spring. Projections from different climate models generally agree that these changes will continue. Likely impacts include increases in near-surface ground temperature, changes in the timing of spring melt-water pulses, and enhanced transportation and agricultural opportunities ( Anisimov et al., 2005a [NPR, MoS, SRC] ). The projected warming also implies a continuation of recent trends toward later freeze-up and earlier break-up of river and lake ice ( Walsh et al., 2005 [NPR, PoC, SRC] ).
Projections of change agree that the retreat of glaciers will continue across Arctic glaciers, with a consequent impact on global sea level ( Meehl et al., 2007 [NPR, ARC, 2007] ). Recent changes in the Greenland ice sheet have, however, been complex. The colder interior has thickened, most probably as a result of recently high precipitation rates, while the coastal zone has been thinning. Thus some studies suggest that overall the ice sheet is growing in thickness ( Krabill et al., 2000 [JoC] ; Johannessen et al., 2005 [JoC] ). However, there is a growing body of evidence for accelerating coastal thinning, a response to recent increases in summer melt ( Abdalati and Steffen, 2001 [JoC] ), and acceleration of many coastal glaciers ( Krabill et al., 2004 [JoC] ; Howat et al., 2005 [JoC] ; Ekstrom et al., 2006 [JoC] ; Luckman et al., 2006 [JoC] ; Rignot and Kanagaratnam, 2006 [JoC] ) suggest that thinning is now dominating the mass balance of the entire ice sheet.
Warming, thawing and decrease in areal extent of terrain underlain by permafrost are expected in response to climatic change in the 21st century ( Sazonova et al., 2004 [SRC] ; Euskirchen et al., 2006 [JoC] ; Lemke et al., 2007 [NPR, ARC, 2007] ). Results from models forced with a range of IPCC climate scenarios indicate that by the mid-21st century the permafrost area in the Northern Hemisphere is likely to decrease by 20 to 35%, largely due to the thawing of permafrost in the southern portions of the sporadic and discontinuous zones, but also due to increasing patchiness in areas that currently have continuous permafrost ( Anisimov and Belolutskaia, 2004 [MoS, SRC] ). Projected changes in the depth of seasonal thawing (base of the active layer) are uniform neither in space nor in time. In the next three decades, active layer depths are likely to be within 10 to 15% of their present values over most of the permafrost area; by the middle of the century, the depth of seasonal thawing may increase on average by 15 to 25%, and by 50% and more in the northernmost locations; and by 2080, it is likely to increase by 30 to 50% and more over all permafrost areas ( Anisimov and Belolutskaia, 2004 [MoS, SRC] ; Instanes et al., 2005 [NPR, SRC] ).
Antarctic
Current and projected changes in the Antarctic ice sheet are discussed in greater detail elsewhere ( Lemke et al., 2007 [NPR, ARC, 2007] ), and are only summarised here. Recent changes in volume of the Antarctic ice sheet are much better mapped and understood than they were in the TAR, but competing theories over the causes still limits confidence in prediction of the future changes. The ice sheet on the Antarctic Peninsula is probably alone in showing a clear response to contemporary climate change (see Section 15.6.3 ), while the larger West Antarctic and East Antarctic ice sheets are showing changes whose attribution to climate change are not clear, but cannot be ruled out. In West Antarctica, there is a suggestion that the dramatic recent thinning of the ice sheet throughout the Amundsen Sea sector is the result of recent ocean change ( Payne et al., 2004 [JoC] ; Shepherd et al., 2004 [JoC] ), but as yet there are too few oceanographic measurements to confirm this interpretation. Indeed, there is evidence that deglaciation of some parts of West Antarctica, as a response to climate change at the end of the last glacial period, is not yet complete ( Stone et al., 2003 [JoC] ). There are still competing theories, but the now clear evidence of ice-sheet change, has reinvigorated debate about whether we should expect a deglaciation of part of the West Antarctic ice sheet on century to millennial timescales ( Vaughan, 2007 [NPR, JoC, SRC, 2007] ). Studies based on satellite observations do not provide unequivocal evidence concerning the mass balance of the East Antarctic ice sheet; some appear to indicate marginal thickening ( Davis et al., 2005 [JoC] ), while others indicate little change ( (Zwally et al., 2005; ) Velicogna and Wahr, 2006 [JoC] ; Wingham et al., 2006 [JoC] ).
Permafrost in ice-free areas, seasonal snow cover, and lake-ice do exist in Antarctica but in such small areas that they are only discussed in respect to particular impacts.
15.4 Key future impacts and vulnerabilities
15.4.1 Freshwater systems and their management
15.4.1.1 Arctic freshwater systems and historical changes
Some freshwater systems exist wholly within the Arctic but many others are fed by river and lake systems further south. The latter includes five of the world’s largest river catchments, which act as major conduits transporting water, heat, sediment, nutrients, contaminants and biota into the Arctic. For these systems, it will be the basin-wide changes that will determine the Arctic impacts.
Historically, the largest changes to northern river systems have been produced by flow regulation, much of it occurring in the headwaters of Arctic rivers. For Canada and Russia, it is these northward-flowing rivers that hold the greatest remaining potential for large-scale hydroelectric development (e.g., Shiklomanov et al., 2000 [NPR, ARC] ; Prowse et al., 2004 [NPR, SRC] ). Similar to some expected effects of climate change, the typical effect of hydroelectric flow regulation is to increase winter flow but also to decrease summer flow and thereby change overall inter-seasonal variability. In the case of the largest Arctic-flowing river in North America, the Mackenzie River, separating the effects of climate change from regulation has proven difficult because of the additional dampening effects on flow produced by natural storage-release effects of major lake systems (e.g., Gibson et al., 2006 [SRC] ; Peters et al., 2006 [SRC] ). For some major Russian rivers (Ob and Yenisei), seasonal effects of hydroelectric regulation have been noted as being primarily responsible for observed trends in winter discharge that were previously thought to be a result of climatic effects ( Yang et al., 2004a [JoC] , b). By contrast, winter flow increases on the Lena River have resulted primarily from increased winter precipitation and warming ( Yang et al., 2002 [JoC] ;( Berezovskaya et al., 2005 ) ). Spatial patterns in timing of flows, however, have not been consistent, with adjacent major Siberian rivers showing both earlier (Lena – Yang et al., 2002 [JoC] ) and later (Yenisei – ( Yang et al., 2004b ) ) spring flows over the last 60 years. Although precipitation changes are often suspected of causing many changes in river runoff, a sparse precipitation monitoring network in the Arctic, makes such linkages very difficult ( Walsh et al., 2005 [NPR, PoC, SRC] ). Seasonal precipitation-runoff responses could be further obscured by the effects of permafrost thaw and related alterations to flow pathways and transfer times ( Serreze et al., 2003 [JoC] ;( Berezovskaya et al., 2005; ) Zhang et al., 2005 [SRC] ).
Over the last half-century, the combined flow from the six largest Eurasian rivers has increased by approximately 7% or an average of 2 km3/yr ( Peterson et al., 2002 [PoC, JoC, ARC] ). The precise controlling factors remain to be identified, but effects of ice-melt from permafrost, forest fires and dam storage have been eliminated as being responsible ( McClelland et al., 2004 [JoC] ). Increased runoff to the Arctic Ocean from circumpolar glaciers, ice caps and ice sheets has also been noted to have occurred in the late 20th century and to be comparable to the increase in combined river inflow from the largest pan-Arctic rivers ( Dyurgerov and Carter, 2004 [ARC] ).
The Arctic contains numerous types of lentic (still-water) systems, ranging from shallow tundra ponds to large lakes. Seasonal shifts in flow, ice cover, precipitation/evapotranspiration and inputs of sediment and nutrients have all been identified as climate-related factors controlling their biodiversity, storage regime and carbon-methane source-sink status ( Wrona et al., 2005 [NPR, SRC] ). A significant number of palaeolimnological records from lakes in the circumpolar Arctic have shown synchronous changes in biological community composition and sedimentological parameters associated with climate-driven regime shifts in increasing mean annual and summer temperatures and corresponding changes in thermal stratification/stability and ice-cover duration (e.g., ( Korhola et al., 2002; ) ( Ruhland et al., 2003; ) Pienitz et al., 2004 [NPR] ; Smol et al., 2005 [PoC, JoC] ; Prowse et al., 2006b [SRC] ).
Permafrost plays a large role in the hydrology of lentic systems, primarily through its influence on substrate permeability and surface ponding of water ( Hinzman et al., 2005 [JoC, SRC] ). Appreciable changes have been observed in lake abundance and area over a 500,000 km2 zone of Siberia during an approximate three-decade period at the end of the last century (see Figure 15.4 Smith et al., 2005 [JoC] ). The spatial pattern of lake disappearance strongly suggests that permafrost thawing is driving the changes.
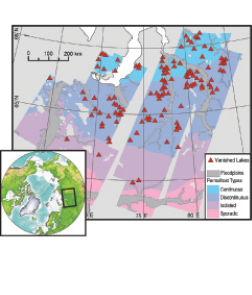
Figure 15.4. Locations of Siberian lakes that have disappeared after
a three-decade period of rising soil and air temperatures (changes registered from satellite imagery from early 1970 s to 1997 - 2004 ), overlaid on various permafrost types. The spatial pattern of lake disappearance suggests that permafrost thawing has driven the observed losses. From Smith et al., 2005 [JoC] . Reprinted with permission from AAAS.
15.4.1.2 Impacts on physical regime
Changes in Arctic freshwater systems will have numerous impacts on the physical regime of the Arctic, particularly affecting hydrological extremes, global feedbacks and contaminant pathways.
Hydrological models based on atmosphere-ocean general circulation models (AOGCMs) have consistently also predicted increases in flow for the major Arctic river systems, with the largest increases during the cold season ( Miller and Russell, 2000 [JoC, MoS] ; Arora and Boer, 2001 [JoC, MoS] ; Mokhov et al., 2003 [MoS] ; Georgievsky et al., 2005 [NPR, ARC] ). Less clear is what may occur during the summer months, with some results suggesting that flow may actually decrease because of evaporation exceeding precipitation (e.g., Walsh et al., 2005 [NPR, PoC, SRC] ). Reductions in summer flow could be enhanced for many watersheds because of increases in evapotranspiration as dominant terrestrial vegetation shifts from non-transpiring tundra lichens to various woody species (e.g., Callaghan et al., 2005 [NPR, SRC] ). CO2-induced reductions in transpiration might offset this, and have been suggested as being responsible for some 20th-century changes in global runoff ( Gedney et al., 2006 [JoC, ARC] ).
Since Arctic river flow is the major component of the freshwater budget of the Arctic Ocean ( Lewis et al., 2000 [NPR, SRC] ), it is important to the supply of freshwater to the North Atlantic and related effects on the thermohaline circulation ( Bindoff et al., 2007 [NPR, 2007] ). The sum of various freshwater inputs to the Arctic Ocean has matched the amount and rate at which freshwater has accumulated in the North Atlantic over the period 1965 to 1995 ( Peterson et al., 2006 [JoC, SRC] ). Under greenhouse gas scenarios, the total annual river inflow to the Arctic Ocean is expected to increase by approximately 10 to 30% by the late 21st century ( Walsh et al., 2005 [NPR, PoC, SRC] ). An additional source of future freshwater input will be from melting of large glaciers and ice caps, most notably from Greenland ( Gregory et al., 2004 [JoC, ARC] ; Dowdeswell, 2006 [JoC] ). The cumulative effect of these increasing freshwater supplies on thermohaline circulation remains unclear but is a critical area of concern ( Loeng et al., 2005 [NPR, ARC] ; Bindoff et al., 2007 [NPR, 2007] ).
Warming is also forecast to cause reductions in river- and lake-ice cover, which will lead to changes in lake thermal structures, quality/quantity of under-ice habitat and effects on ice jamming and related flooding ( Prowse et al., 2006a [SRC] ). Specific to the latter, forecasts of earlier snowmelt freshets could create conditions more conducive to severe break-up events ( Prowse and Beltaos, 2002 [SRC] ), although a longer period of warming could also reduce severity ( (Smith, 2000 ) ). This effect, however, is likely to be offset on some large northward-flowing rivers because of reduced regional contrasts in south-to-north temperatures and related hydrological and physical gradients ( Prowse et al., 2006a [SRC] ).
Projected changes of permafrost, vegetation and river-runoff may have noticeable impacts on river morphology, acting through destabilisation of banks and slopes, increased erosion and sediment supply, and ultimately leading to the transformation between multi- and single-channel types. Geological reconstructions and numerical simulations indicate that such transformations and also erosion events and flood risks occur especially at times of permafrost degradation ( Bogaart and van Balen, 2000 [MoS] ; Vandenberghe, 2002 [SRC] ). Such changes are largely controlled by thresholds in sediment supply to the river and discharge ( Vandenberghe, 2001 [SRC] ). However, historical examples have shown that variability in flow regime is less important than variability in sediment supply, which is especially determined by the vegetation cover ( (Huisink et al., 2002; ) Vandenberghe and Huisink, 2003 [NPR, SRC] ). Thus an increasingly denser vegetation cover may counter increased sediment discharge, which has been modelled to rise in Arctic rivers with both increases in air temperature and water discharge ( Syvitski, 2002 [MoS] ).
Various changes in Arctic hydrology have the potential to effect large changes in the proportion of pollutants (e.g., persistent organic pollutants and mercury) that enter Arctic aquatic systems, either by solvent-switching or solvent-depleting processes (e.g., MacDonald et al., 2003 [NPR] ). Given that the Arctic is predicted to be generally ‘wetter’, the increase in loadings of particulates and contaminants that partition strongly into water might more than offset the reductions expected to accrue from reductions in global emissions (e.g., MacDonald et al., 2003 [NPR] ). Shifts in other hydrological regime components such as vegetation, runoff patterns and thermokarst drainage ( Hinzman et al., 2005 [JoC, SRC] ) all have the capacity to increase contaminant capture. Changes in aquatic trophic structure and related rate functions (see Section 15.4.1.3 ) have further potential to alter the accumulation of bio-magnifying chemicals within food webs.
15.4.1.3 Impacts on aquatic productivity and biodiversity
Projected changes in runoff, river- and lake-ice regimes, and seasonal and interannual water balance and thermal characteristics will alter biodiversity and productivity relationships in aquatic ecosystems ( Walsh et al., 2005 [NPR, PoC, SRC] ; Prowse et al., 2006b [SRC] ; Wrona et al., 2006a [SRC] ). Ultimately the dispersal and geographical distribution patterns of aquatic species will be altered, particularly for fish ( Reist et al., 2006a [SRC] ). Extension of the ice-free season may lead to a decline in fish habitat availability and suitability, particularly affecting species such as lake trout (Salvelinus namaycush) that prefer colder waters ( (Hobbie et al., 1999; ) Reist et al., 2006b [SRC] ). The projected enhanced river flows will also increase sediment transport and nutrient loading into the Arctic Ocean, thereby affecting estuarine and marine productivity ( (Carmack and Macdonald, 2002 ) ).
Increased permafrost thawing and deepening of the active layer will increase nutrient, sediment and carbon loadings, enhancing microbial and higher trophic level productivity in nutrient-limited systems. As water-column dissolved organic carbon (DOC) concentration increases, penetration of damaging UV radiation and photochemical processing of organic material would decline, although not as prominently in highly productive systems ( Reist et al., 2006b [SRC] ; Wrona et al., 2006a [SRC] ). Enhanced sediment loadings will negatively affect benthic and fish-spawning habitats by increasing the biological oxygen demand and hypoxia/anoxia associated with sedimentation, and contribute to habitat loss through infilling ( Reist et al., 2006a [SRC] ). Whether freshwater systems will function as net carbon sinks or sources depends on the complex interactions between temperature, nutrient status and water levels ( Frey and Smith, 2005 [JoC] ; Flanagan et al., 2006 [JoC, SRC] ). Initial permafrost thaw will form depressions for new wetlands and ponds interconnected by new drainage networks. This will allow for the dispersal and establishment of new aquatic communities in areas formerly dominated by terrestrial species ( Wrona et al., 2006b [SRC] ). As the permafrost thaws further, surface waters will increasingly drain into groundwater systems, leading to losses in freshwater habitat.
Southerly species presently limited by temperature/ productivity constraints will probably colonise Arctic areas, resulting in new assemblages. Many of these, particularly fishes, will be likely to out-compete or prey upon established Arctic species, resulting in negative local effects on these ( Reist et al., 2006a [SRC] ). These southern emigrants to the Arctic will also bring with them new parasites and/or diseases to which Arctic species are not adapted, thereby increasing mortality ( Wrona et al., 2006b [SRC] ). Direct environmental change combined with indirect ecosystem shifts will significantly impact local faunas by reducing productivity, abundance and biodiversity. Such effects will be most severe for freshwater fish that rely entirely upon local aquatic ecosystems ( Reist et al., 2006c [SRC] ). Distributions of anadromous fish, which migrate up rivers from the sea to breed in freshwater, will probably shift as oceanic conditions and freshwater drainage patterns are affected ( Reist et al., 2006c [SRC] ); as will the geographical patterns of habitat use of migratory aquatic birds and mammals ( Wrona et al., 2005 [NPR, SRC] ). Important northern fish species such as broad whitefish (Coregonus nasus), Arctic char (Salvelinus alpinus), inconnu (Stenodus leucichthys), Arctic grayling (Thymallus arcticus) and Arctic cisco (Coregonus autumnalis) will probably experience population reductions and extirpations (e.g., due to reproductive failures), contraction of geographical ranges in response to habitat impacts, and competition and predation from colonising species ( Reist et al., 2006a [SRC] , b, c).
15.4.1.4 Impacts on resource use and traditional economies/livelihoods
Given the large hydrological changes expected for Arctic rivers, particularly regarding the magnitude of the spring freshet, climate-induced changes must be factored into the design, maintenance and safety of existing and future development structures (e.g., oil and gas drilling platforms, pipelines, mine tailings ponds, dams and impoundments for hydroelectric production) (World Commission on Dams, 2000; Prowse et al., 2004 [NPR, SRC] ; Instanes et al., 2005 [NPR, SRC] ).
Freshwater sources are critical to human health, especially for many northern communities that rely on surface and/or groundwater, often untreated, for drinking water and domestic use (United States Environmental Protection Agency, 1997; Martin et al., 2005 [NPR, SRC] ). Direct use of untreated water from lakes, rivers and large pieces of multi-year sea ice is considered to be a traditional practice, despite the fact that it poses a risk to human health via the transmission of water-borne diseases (e.g., Martin et al., 2005 [NPR, SRC] ). Such risks may increase with changes in migration and northward movement of species and their related diseases. Changes in hydrology may also decrease the availability and quality of drinking water, particularly for coastal communities affected by rising sea levels where sea-water contamination could affect groundwater reserves ( (Warren et al., 2005 ) ).
Northern freshwater ecosystems provide many services to Arctic peoples, particularly in the form of harvestable biota used to support both subsistence and commercial economies ( Reist et al., 2006b [SRC] ). Shifts in ecosystem structure and function will result in substantial changes in the abundance, replenishment, availability and accessibility of such resources which, in turn, will alter local resource use and traditional and subsistence lifestyles ( Nuttall et al., 2005 [NPR, SRC] ; Reist et al., 2006b [SRC] ). It is unlikely that such changes related to natural freshwater systems would be offset by increased opportunity for freshwater aquaculture resulting from a warming climate. Thus, conservation of Arctic aquatic biodiversity, maintenance of traditional and subsistence lifestyles, and continued viability and sustainable use of Arctic freshwater resources will present significant challenges for Arctic peoples, resource managers and policy-makers ( Wrona et al., 2005 [NPR, SRC] ; Reist et al., 2006b [SRC] ).
15.4.2 Terrestrial ecosystems and their services
15.4.2.1 Historical and current changes in Arctic terrestrial ecosystems
Climatic changes during the past 20,000 years and more have shaped current biodiversity, ecosystem extent, structure and function. Arctic species diversity is currently low, partly because of past extinction events (FAUNMAP Working Group, 1996 ). As a group, large mammals are in general more vulnerable to current change than in the past when the group contained many more species. Also, tundra ecosystem extent, particularly in Eurasia, is now less than during the glacial period, when extensive tundra-steppe ecosystems existed ( Callaghan et al., 2005 [NPR, SRC] ). Modern habitat fragmentation (e.g., Nellemann et al., 2001 [NPR] ), stratospheric ozone depletion, and spread of contaminants, compound the ongoing impacts of anthropogenic climate change and natural variability on ecosystems and their services.
Traditional ecological knowledge (TEK, see Section 15.6.1 ) from Canada has recorded current ecosystem changes such as poor vegetation growth in eastern regions associated with warmer and drier summers; increased plant biomass and growth in western regions associated with warmer, wetter and longer summers; the spreading of some existing species, and new sightings of a few southern species; and changing grazing behaviours of musk oxen and caribou as the availability of forage increases in some areas ( (Riedlinger and Berkes, 2001; ) Thorpe et al., 2001 [NPR] ; Krupnik and Jolly, 2002 [NPR] ).
In northern Fennoscandia the cycles of voles, and possibly also of lemmings, have become considerably dampened since the 1980 s, due to low spring peak densities, and it is likely that these changes are linked to poorer winter survival due to changing snow conditions ( (Yoccoz and Ims, 1999; ) Henttonen and Wallgren, 2001 [NPR] ;( Ims and Fuglei, 2005 ) ). Arctic fox, lesser white fronted goose and shore lark have declined dramatically ( (Elmhagen et al., 2000 ) ) and moose, red fox and some southern bird species have spread northwards ( (Hörnberg, 1995; ) ( Tannerfeldt et al., 2002 ) ), although the specific role of climate change is unknown. Some migrant bird populations, particularly Arctic waders, have declined substantially ( (Stroud et al., 2004 ) ) due to various causes including climate change and loss of habitat on migration routes and wintering grounds ( (Morrison et al., 2001, ) 2004; ( Zöckler, 2005 ) ). In contrast, populations of Arctic breeding geese, which in winter increasingly feed on agricultural crops or unharvested grain, have shown a geometric increase in numbers in Europe and North America, which has led to intense foraging in Arctic coastal breeding habitats, loss of vegetation and the occurrence of hypersaline soils ( Jefferies et al., 2006 [NPR, SRC] ). Although there are examples from temperate latitudes, evidence for early arrival of migratory birds in the Arctic is weak ( Gauthier et al., 2005 [JoC] ), emphasising the need for adequate monitoring programmes ( Both et al., 2005 [NPR] ). Some populations of caribou/reindeer, which are essential to the culture and subsistence of several Arctic peoples, are currently in decline ( Russell et al., 2002 [NPR] ; Chapin et al., 2005a [NPR, SRC] ), mainly due to social and cultural factors. However, climate impacts have also affected some populations. Icing events during warmer winters that restrict access to frozen vegetation have impacted some reindeer/caribou populations and high-Arctic musk oxen populations ( (Forchhammer and Boertmann, 1993; ) ( Aanes et al., 2000; ) Callaghan et al., 2005 [NPR, SRC] and references therein).
Evidence of recent vegetation change is compelling. Aerial photographs show increased shrub abundance in Alaska in 70% of 200 locations ( Sturm et al., 2001 [JoC] ; Tape et al., 2006 [JoC] ). Along the Arctic to sub-Arctic boundary, the tree line has moved about 10 km northwards, and 2% of Alaskan tundra on the Seward Peninsula has been displaced by forest in the past 50 years ( Lloyd et al., 2003 [JoC] ). In some areas, the altitude of the tree line has risen, for example by about 60 m in the 20th century in sub-Arctic Sweden ( Callaghan et al., 2004 [MoS, SRC] ; Truong et al., 2007 [2007] ), although the tree line has been stable or become lower in other localities ( (Dalen and Hofgaard, 2005 ) ). Bog growth has caused tree death in parts of the Russian European Arctic ( (Crawford et al., 2003 ) ). The pattern of northward and upward tree-line advances is comparable with earlier Holocene changes ( MacDonald et al., 2000 [ARC] ; Esper and Schweingruber, 2004 [JoC] ) (see below for rates of advance). In addition to changes in woody vegetation, dry-habitat vegetation in sub-Arctic Sweden has been partly displaced by wet-habitat vegetation because of permafrost degradation in the discontinuous permafrost zone ( Christensen et al., 2004 [JoC] ; Malmer et al., 2005 [JoC] ). Similarly, in northern Canada, up to 50% of peat plateau permafrost has thawed at four sites in the discontinuous permafrost zone ( Beilman and Robinson, 2003 [NPR] ).
There is also recent evidence of changes in growing season duration and timing, together with plant productivity, but patterns are spatially variable. Analyses of satellite images indicate that the length of growing season is increasing by 3 days per decade in Alaska and 1 day per decade in northern Eurasia ( (McDonald et al., 2004; ) Smith et al., 2004 [Ambiguous] ; McGuire et al., 2007 [NPR, SRC, 2007] ), but there has been a delayed onset of the growing season in the Kola Peninsula during climatic cooling over the past two decades ( Høgda et al., 2007 [NPR, 2007] ). Remote sensing estimates of primary productivity also show spatial variability: there were increases in the southern Arctic and decreases in the central and eastern Russian Arctic between 1982 and 1999 ( Nemani et al., 2003 [JoC, ARC] ).
15.4.2.2 Projected changes in biodiversity, vegetation zones and productivity
Where soils are adequate for forest expansion, species richness will increase as relatively species-rich forest displaces tundra (see Figure 15.3 Callaghan et al., 2005 [NPR, SRC] ). Some species in isolated favourable microenvironments far north of their main distribution are very likely to spread rapidly during warming. Except for the northernmost and highest-Arctic species, species will generally extend their ranges northwards and higher in altitude, while the dominance and abundance of many will decrease. Likely rates of advance are uncertain; although tree-line advance of up to 25 km/yr during the early Holocene have been recorded, rates of 2 km/yr and less are more probable ( (Payette et al., 2002; ) Callaghan et al., 2005 [NPR, SRC] ). Trophic structure is relatively simple in the Arctic, and decreases in the abundance of keystone species are expected to lead to ecological cascades, i.e., knock-on effects for predators, food sources etc. Local changes in distribution and abundance of genetically different populations will be the initial response of genetically diverse species to warming ( (Crawford, 2004 ) ). Arctic animals are likely to be most vulnerable to warming-induced drying (invertebrates); changes in snow cover and freeze-thaw cycles that affect access to food and protection from predators; changes that affect the timing of behaviour (e.g., migration and reproduction); and influx of new competitors, predators, parasites and diseases. Southern species constantly reach the Arctic but few become established ( Chernov and Matveyeva, 1997 [NPR] ). As a result of projected climate change, establishment will increase and some species, such as the North American mink, will become invasive, while existing populations of weedy southern plant species that have already colonised some Arctic areas are likely to expand. The timing of bird migrations and migration routes are likely to change as appropriate Arctic habitats become less available ( Callaghan et al., 2005 [NPR, SRC] ; Usher et al., 2005 [NPR, SRC] ).
Warming experiments that adequately reproduced natural summer warming impacts on ecosystems across the Arctic showed that plant communities responded rapidly to 1-3°C warming after two growing seasons, that shrub growth increased as observed under natural climate warming, and that species diversity decreased initially ( Walker et al., 2006 [JoC, SRC] ). Experimental warming and nutrient addition showed that mosses and lichens became less abundant when vascular plants increased their growth ( Cornelissen et al., 2001 [SRC] ; van Wijk et al., 2003 [JoC, SRC] ). CO2-enrichment produced transient plant responses, but microbial communities changed in structure and function ( Johnson et al., 2002 [JoC, SRC] ) and frost hardiness of some plants decreased ( Beerling et al., 2001 [MoS, SRC] ) making them more susceptible to early frosts. Supplemental ultraviolet-B caused few plant responses but did reduce nutrient cycling processes ( Callaghan et al., 2005 [NPR, SRC] ; Rinnan et al., 2005 [JoC] ). Such reductions could potentially reduce plant growth.
A ‘moderate’ projection for 2100 for the replacement of tundra areas by forest is about 10% ( Sitch et al., 2003 [JoC, MoS, ARC] ; see Figure 15.3 ), but estimates of up to 50% have also been published ( White et al., 2000 [JoC, MoS] ). However, impacts of changing hydrology, active layer depth and land use are excluded from these models. These impacts can be large: for example, ( Vlassova 2002 ) ) suggests that 475,000 km2 of tree-line forest has been destroyed in Russia, thereby creating tundra-like ecosystems. Narrow coastal strips of tundra (e.g., in parts of the Russian European Arctic) will be completely displaced as forest reaches the Arctic Ocean. During 1960 to 2080, tundra is projected to replace about 15 to 25% of the polar desert, and net primary production (NPP) will increase by about 70% (2.8 to 4.9 Gt of carbon) ( Sitch et al., 2003 [JoC, MoS, ARC] ). Geographical constraints on vegetation relocation result in large sub-regional variations in projected increases of NPP, from about 45% in fragmented landmasses to about 145% in extensive tundra areas ( Callaghan et al., 2005 [NPR, SRC] ).
Climate warming is likely to increase the incidence of pests, parasites and diseases such as musk ox lung-worm ( (Kutz et al., 2002 ) ) and abomasal nematodes of reindeer ( (Albon et al., 2002 ) ). Large-scale forest fires and outbreaks of tree-killing insects that are triggered by warm weather are characteristic of the boreal forest and are likely to increase in extent and frequency ( Juday et al., 2005 [NPR] ), creating new areas of forest tundra. During the 1990 s, the Kenai Peninsula of south-central Alaska experienced a massive outbreak of spruce bark beetle over 16,000 km2 with 10-20% tree mortality ( Juday et al., 2005 [NPR] ). Also following recent climate warming, spruce budworm has reproduced further north, reaching problematic numbers in Alaska ( Juday et al., 2005 [NPR] ), while autumn moth defoliation of mountain birch trees, associated with warm winters in northern Fennoscandia, has occurred over wide areas and is projected to increase ( Callaghan et al., 2005 [NPR, SRC] ).
15.4.2.3 Consequences of changes in ecosystem structure and function for feedbacks to the climate system
Climate warming will decrease the reflectivity of the land surface due to expansion of shrubs and trees into tundra ( Eugster et al., 2000 [JoC] ); this could influence regional ( Chapin et al., 2005a [NPR, SRC] ) and global climate ( Bonan et al., 1992 [JoC] ;( Thomas and Rowntree, 1992; ) Foley et al., 1994 [JoC] ; Sturm et al., 2005 [JoC] ; McGuire et al., 2007 [NPR, SRC, 2007] ).
Measurements show great spatial variability in the magnitude of sink (net uptake) or source (net release) status for carbon, with no overall trend for the Arctic ( Corradi et al., 2005 [JoC] ). In contrast, models suggest that overall the Arctic is currently a small sink of about 20 ± 40 g carbon m2/yr ( McGuire et al., 2000 [JoC, MoS] ; Sitch et al., 2003 [JoC, MoS, ARC] , 2007 ). The high uncertainties in both measurements and model projections indicate that the Arctic could be either a sink or source of carbon. Thus, currently circumpolar Arctic vegetation and the active layer are unlikely to be a large source or sink of carbon in the form of CO2 ( Callaghan et al., 2005 [NPR, SRC] ; Chapin et al., 2005a [NPR, SRC] ). They are, however, most probably a source of positive radiative forcing due to large methane emissions; even in tundra areas that are net sinks of carbon, significant emissions of methane lead to positive forcing ( Friborg et al., 2003 [JoC] ; Callaghan et al., 2005 [NPR, SRC] ).
Higher temperatures, longer growing seasons and projected northward movement of productive vegetation are likely to increase photosynthetic carbon capture in the longer term, whereas soil warming is likely to increase trace gas emissions in the short term. Drying or wetting of tundra concurrent with warming and increased active-layer depth (see Section 15.3.4 ) will determine the magnitude of carbon fluxes and the balance of trace gases that are involved. Drying has increased sources in Alaska ( Oechel et al., 2000 [JoC] ), whereas wetting has increased sinks in Scandinavian and Siberian peatlands ( Aurela et al., 2002 [JoC] ; Johansson et al., 2006 [JoC, MoS] ).
Models project that the Arctic and sub-Arctic are likely to become a weak sink of carbon during future warming (an increase in carbon storage in vegetation, litter and soil of about 18.3 Gt carbon between 1960 and 2080 ), although there is high uncertainty ( Sitch et al., 2003 [JoC, MoS, ARC] ; Callaghan et al., 2005 [NPR, SRC] ). Increased carbon emissions from projected increases in disturbances and land use, and net radiative forcing resulting from the changing balance between methane and carbon dioxide emissions ( Friborg et al., 2003 [JoC] ; Johansson et al., 2006 [JoC, MoS] ) are particular uncertainties. Wetting, from increased precipitation and permafrost thawing, is projected to increase fluxes of methane relative to carbon dioxide from the active layer and thawing permafrost ( Walter et al., 2006 [JoC] ).
Changes in forest area will lead to both negative and positive feedbacks on climate. According to one coupled climate model, the negative feedback of carbon sequestration and the positive feedback of reduced albedo interact. This model predicts that the central Canadian boreal forests will give net negative feedback through dominance of increased carbon sequestration, while in the forests of Arctic Russia decreased albedo will dominate, giving net positive feedback ( Betts and Ball, 1997 [JoC] ; Betts, 2000 [JoC, ARC] ).
15.4.2.4 Impacts on resource use, traditional economies and lifestyles
Terrestrial resources are critical aspects of Arctic residents’ livelihoods, culture, traditions and health (Arctic Monitoring and Assessment Programme, 2003; Chapin et al., 2005a [NPR, SRC] ). Per capita consumption of wild foods by rural Alaskans is 465 g per day (16% land mammals, 10% plant products) and consumption by urban Alaskans is 60 g per day. The collective value of these foods in the state is estimated at about US$200 million/yr. Consumption in Canadian Arctic communities ranges from 106 g per day to 440 g per day, accounting for 6 to 40% of total energy intake and 7 to 10% of the total household income in Nunavik and Nunavut ( (Kuhnlein et al., 2001; ) ( Chabot, 2004 ) ). Terrestrial ecosystem resources include caribou/reindeer, moose, musk ox, migratory birds and their eggs, and plants and berries (Arctic Monitoring and Assessment Programme, 2003; Chapin et al., 2005a [NPR, SRC] ). Wild and domesticated caribou/reindeer are particularly important, as they provide food, shelter, clothing, tools, transportation and, in some cases, marketable goods ( Klein, 1989 [NPR] ; Paine, 1994 [NPR] ;( Kofinas et al., 2000; ) Jernsletten and Klokov, 2002 [NPR] ). Wood, sods, peat and coal are used locally as fuels throughout the north. Despite the significant role these resources represent to Arctic residents, ties to subsistence activities among indigenous peoples are deteriorating because of changes in lifestyles, cultural, social, economic and political factors ( Chapin et al., 2005a [NPR, SRC] ). These ties are expected to continue decreasing as climate-driven changes in terrestrial ecosystems influence conditions for hunting, decreases in natural resources, and loss of traditional knowledge. Together these shifts are turning previously well-adapted Arctic peoples into “strangers in their own lands” ( Berkes, 2002 [NPR] ).
Agriculture in southern parts of the Arctic is limited by short, cool growing seasons and lack of infrastructure, including limited local markets because of small populations and long distances to large markets ( Juday et al., 2005 [NPR] ). The northern limit of agriculture may be roughly approximated by a metric based on the cumulative degree-days above +10°C ( Sirotenko et al., 1997 [JoC] ). By the mid-21st century, climatic warming may see displacement of its position to the north by a few hundred kilometres over most of Siberia, and up to 100 km elsewhere in Russia ( Anisimov and Belolutskaia, 2001 [MoS, SRC] ). Thus climate warming is likely to lead to the opportunity for an expansion of agriculture and forestry where markets and infrastructure exist or are developed. While conservation management and protected areas are extensive in the Arctic, these only protect against direct human actions, not against climate-induced vegetation zone shifts, and decisions need to be made about the goals and methods of conservation in the future ( Callaghan et al., 2005 [NPR, SRC] ; Klein et al., 2005 [NPR] ; Usher et al., 2005 [NPR, SRC] ).
15.4.3 Marine ecosystems and their services in the Arctic
15.4.3.1 Historical changes in marine ecosystems
Water temperatures in the North Atlantic have fluctuated over the last 200 years. The effects of these temperature variations have been profound, impacting plankton communities as well as larval drift, distribution and abundance of many fish stocks including southern invaders and, especially, the commercially important cod and herring ( Loeng et al., 2005 [NPR, ARC] ; Vilhjálmsson et al., 2005 [NPR, SRC] ; see Section 15.6.3 ). These climatic impacts on fish stocks are superimposed on those arising from exploitation.
15.4.3.2 Likely general effects of a warming ocean climate
Changing climatic conditions in Arctic and sub-Arctic oceans are driving changes in the biodiversity, distribution and productivity of marine biota, most obviously through the reduction of sea ice. As the sea-ice edge moves northward, the distribution of crustaceans (copepods and amphipods), adapted for life at the sea-ice edge, and fish such as polar cod (Boreogadus saida), which forage on them, will shift accordingly and their abundance diminish ( (Sakshaug et al., 1994 ) ). This reduction is likely to seriously impact other predators, e.g., seals, sea birds and polar bears (Ursus maritimus), dependent on sea ice for feeding and breeding (see Chapter 4 , Box 4.3 ( Sakshaug et al., 1994 ) ) as well as humans depending on them ( Loeng et al., 2005 [NPR, ARC] ; Vilhjálmsson et al., 2005 [NPR, SRC] ).
Thinning and reduced coverage of Arctic sea ice are likely to substantially alter ecosystems that are in close association with sea ice ( Loeng et al., 2005 [NPR, ARC] ). Polar cod, an important member of these ecosystems, is a prime food source for many marine mammals. Ringed seals, which are dependent on sea ice for breeding, moulting and resting, feed on ice amphipods and cod. Premature break-up of ice may not only lead to high mortality of seal pups but also produce behavioural changes in seal populations ( Loeng et al., 2005 [NPR, ARC] ). Polar bears, a top predator, are highly dependent on both sea ice and ringed seals (see Chapter 4 , Box 4.3 ). Initially, the loss of sea ice and the subsequent deleterious effects are likely to occur at the southern distribution limit of polar bears, where early melt and late freezing of sea ice extend the period when the bears are restricted to land and only limited feeding can occur. Recently, the condition of adult bears has declined in the Hudson Bay region and first-year cubs come ashore in poor condition ( Stirling et al., 1999 [ARC] ; Derocher et al., 2004 [ARC] ;( Stirling and Parkinson, 2006 ) ). As a proportion of the population, the number of cubs has fallen as a result of the early break-up of sea ice. Loss of sea ice may also adversely affect other Arctic marine mammals, such as walrus (Odobenus rosmarus) that use sea ice as a resting platform and which occupy a narrow ecological range with restricted mobility. Similarly, the narwhal (Monodon monoceros) and the bowhead whale (Balaena mysticetus) are dependent on sea-ice organisms for feeding and polynyas for breathing ( Loeng et al., 2005 [NPR, ARC] ). The early melting of sea ice may lead to an increasing mismatch in the timing of these sea-ice organisms and secondary production that severely affects the populations of sea mammals ( Loeng et al., 2005 [NPR, ARC] ).
However, with an increase in open water, primary and secondary production south of the ice edge will increase and this will benefit almost all of the most important commercial fish stocks in Arctic and sub-Arctic seas; for example, cod (Gadus morhua) and herring (Clupea harengus) in the North Atlantic and walleye pollock (Theragra chalcogramma) in the Bering Sea; species that currently comprise about 70% of the total catch in these areas. However, some coldwater species (e.g., northern shrimp (Pandalus borealis) and king crab (Paralithoides spp.) may lose habitat ( Vilhjálmsson et al., 2005 [NPR, SRC] ).
15.4.3.3 Predicting future yields of commercial and forage stocks
Quantitative predictions of the responses of commercial and forage fish stocks to changes in ocean temperature are very difficult to make because (a) resolution of existing predictive models is insufficient, (b) exploitation has already altered stock sizes and basic biology/ecology, so stocks cannot be expected to react as they did historically, and (c) the effect of a moderate climate change (+1-3°C) may well be of less importance than sound fisheries policies and their effective enforcement. The examples given in Box 15.1 show how such climate change, exploitation and other factors affecting marine ecosystems may interact strongly at the northern extreme of the range of a species, while having less effect on the same species further south (e.g., Rose et al., 2000 [SRC] ; Rose, 2004 [SRC] ; Drinkwater, 2005 [MoS] ).
Box 15.1. Atlantic cod in the 20th century: historical examples
This box illustrates how cod populations have responded through the 20th century and until 2005 to the multiple stresses of ocean temperature change, exploitation, changing abundance of food species and predators. All four cod populations show substantial decreases to the present, and suggest that they are now vulnerable to future changes in both climate and exploitation. However, the data suggest that the vulnerability of stocks south of the polar front is less than that to the north of the polar front.
Greenland
Ocean temperature records, begun off West Greenland in the 1870 s, showed very cold conditions until temperatures warmed suddenly around 1920, and maintained high levels until they dropped suddenly in the late 1960 s ( (Jensen, 1939; ) ( Buch et al., 1994; ) Vilhjálmsson, 1997 [SRC] ; see Figure 15.5 a). There were no Atlantic cod in Greenland waters in the latter half of the 19th century ( (Jensen, 1926; ) ( Buch et al., 1994 ) ), while there was a good cod fishery off Iceland ( (Jensen, 1926, ) 1939; ( Buch et al., 1994 ) ). Concurrent with the warming in the early 1920 s, large numbers of juvenile cod drifted from Iceland to West Greenland and started a self-supporting stock there, which vanished in the 1970 s ( (Buch et al., 1994; ) Vilhjálmsson, 1997 [SRC] ; Vilhjálmsson et al., 2005 [NPR, SRC] ).
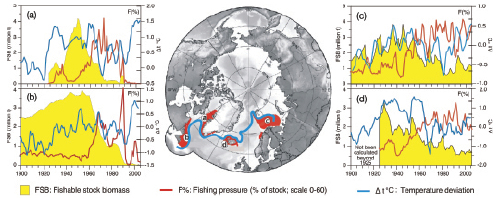
Figure 15.5. The geographical distribution of four major cod stocks in the North Atlantic (red patches). The continuous blue line indicates an average geographical position of the Polar Front. The graphs (a: West Greenland; b: Newfoundland/Labrador; c: Barents Sea, and d: Iceland) show the developments of fishable stock (yellow shading), catches (red line) and temperature (blue line) during the period 1900 - 2005 . Data sources: Greenland ( (Buch et al., 1994; ) ( Horsted, 2000; ) ICES, 2006 [NPR] ); Newfoundland/Labrador ( Harris, 1990 [NPR] ; Lilly et al., 2003 [NPR] ); Iceland ( (Schopka, 1994; ) Hafrannsóknastofnunin, 2006; ICES, 2006 [NPR] ); Barents Sea( (Bochkov, 1982; ) Hylen, 2002 [NPR] ; ICES, 2005a [NPR] ), data since 1981 were kindly provided by the Polar Research Institute of Marine Fisheries and Oceanography (PINRO), Murmansk, Russia.
Comparison of catches and temperature records shows that the occurrence of cod off Greenland depends principally on West Greenland water temperature ( (Horsted, 2000 ) ). Thus the reappearance of cod off Greenland will probably depend on drift of juvenile cod from Iceland as it did in the 1920 s. West Greenland waters may now be sufficiently warm for this to happen. Indeed such drifts did occur in 2003 and 2005, but the numbers were small; a probable consequence of the depleted and younger spawning stock of Icelandic cod, which has not produced a strong year class for 20 years ( ICES, 2005b [NPR] ).
Newfoundland/Labrador
Beginning in the 16th century, annual catches from this stock increased until the mid- 1800 s. From 1920 to 1960, catches varied then increased rapidly, peaking in 1968 . Catches then dropped sharply until about 1977 when Canada acquired its 200-mile Exclusive Economic Zone ( Rose, 2004 [SRC] ). Total allowable catches were increased in the mid- to late 1980 s, but dropped again after 1989 . A moratorium on fishing was imposed in 1992 (see Figure 15.5 b). While fishing was the primary cause of the decline ( (Walters and Maguire, 1996 ) ), decreased productivity in cod and a reduction in their primary food (capelin) also occurred in the late 1980 s and 1990 s ( Rose and O’Driscoll, 2002 [SRC] ;( Shelton et al., 2006 ) ). Furthermore, a moratorium on seal hunting led to an explosion in seal abundance and thus increased mortalities of their prey, including cod ( Lilly et al., 2003 [NPR] ). High mortality remains the key obstacle to population growth. While future warming climate is likely to promote recovery of cod ( Drinkwater, 2005 [MoS] ), an increase in abundance of the main forage fish, capelin, is likely to be a necessary precursor.
Iceland/Barents Sea
In the face of exploitation, the comparative resilience of these cod stocks is high because they are south of the polar front and, therefore, well inside the limit tolerance of the species to cold temperatures (see Figure 15.5 c, d).
Changes in distribution of commercial stocks may lead to conflicts over fishing rights and will require effective negotiations to generate solutions regarding international co-operation in fisheries management ( Vilhjálmsson et al., 2005 [NPR, SRC] ).
In addition, rising water temperatures will lead to an increased risk of harmful algal blooms and occurrences of other marine pests and pollution, hazards that will be multiplied by increased shipping ( Loeng et al., 2005 [NPR, ARC] ; Vilhjálmsson et al., 2005 [NPR, SRC] ).
15.4.4 Marine ecosystems and their services in the Antarctic
Southern Ocean ecosystems are far from pristine. Over the last 200 years some seal and whale species (e.g., Antarctic fur seals, blue and fin whales) were exploited almost to extinction, then fisheries developed. From the 1960 s, fin-fish were exploited, and in the Scotia Sea and surrounding areas stocks of these fish were reduced to low levels and have not yet recovered. In contrast to the Arctic, however, the management of Southern Ocean fisheries is based on an ecosystem approach, within an international convention. The Convention on the Conservation of Antarctic Marine Living Resources (CCAMLR), part of the Antarctic Treaty, was designed to maintain the natural marine ecosystem while allowing sustainable exploitation, and emphasises the need to consider the wider context of the exploitation of individual species, taking account of the entire food web and environmental variations. The CCAMLR applies to areas south of the Antarctic polar front, and management decisions are made by consensus of the Member States ( (Constable et al., 2000 ) ).
The current major fin-fish fishery is for the Patagonian toothfish (Dissostichus eleginoides), and to a lesser extent for the mackerel icefish (Champsocephalus gunnari). The fishery for Antarctic krill (Euphausia superba) developed during the 1970 s, peaked in the 1980 s at over 500,000 t/yr and now operates at about 100,000 t/yr ( (Jones and Ramm, 2004 ) ), a catch that is well below the precautionary limits set within CCAMLR for maintaining the stock.
During the 20th century there were significant changes in air temperatures, sea-ice and ocean temperatures around the Antarctic Peninsula (see Section 15.6.3 ) and in the Scotia Sea. Over 50% of the krill stock was lost in the Scotia Sea region ( Atkinson et al., 2004 [JoC] ), which is the major area for krill fishing. The decline in the abundance of krill in this area appears to be associated with changes in sea ice in the southern Scotia Sea and around the Antarctic Peninsula ( Atkinson et al., 2004 [JoC] ). Future reductions in sea ice may therefore lead to further changes in distribution and abundance across the whole area, with consequent impacts on food webs where krill are currently key prey items for many predator species and where krill fishing occurs.
For other species the uncertainty in climate predictions leads to uncertainty in projections of impacts, but increases in temperatures and reductions in winter sea ice would undoubtedly affect the reproduction, growth and development of fish and krill, leading to further reductions in population sizes and changes in distributions. However, the potential for species to adapt is mixed, some ‘cold-blooded’ (poikilothermic) organisms may die if water temperatures rise to 5-10°C ( Peck, 2005 [NPR] ), while the bald rock cod (Pagothenia borchgrevinki), which uses the specialisation of anti-freeze proteins in its blood to live at sub-zero temperatures, can acclimatise so that its swimming performance at +10°C is similar to that at -1°C ( (Seebacher et al., 2005 ) ).
The importance of ocean transport for connecting Southern Ocean ecosystems has been increasingly recognised. Simple warming scenarios may indicate that exploitation effects would be shifted south, but it is also likely that other species may become the target of new fisheries in the same areas. More complex changes in patterns of ocean circulation could have profound effects on ocean ecosystems and fisheries, although not all changes may be negative and some species may benefit. Complex interactions in food webs may, however, generate secondary responses that are difficult to predict. For example, reductions in krill abundance may have negative effects on species of fish, as they become a greater target for predators. Importantly, the impact of changes in these ecosystems will not be confined to the Southern Ocean. Many higher predator species depend on lower-latitude systems during the Antarctic winter or the breeding seasons.
The fundamental precautionary basis for managing exploitation in a changing environment is in place in CCAMLR, but longer duration and more spatially extensive monitoring data are required in order to help identify change and its effects.
15.4.5 Human health and well-being
The impact of projected climate change on the diverse communities of the Arctic can only be understood in the context of the interconnected social, cultural, political and economic forces acting on them ( Berner et al., 2005 [NPR, SRC] ). However, such impacts on the health and well-being of Arctic residents are, and will be, tangible and ongoing. Recently, significant research has been conducted on the health and well-being of indigenous populations in the Arctic and the role of environmental change as a determinant of health; accordingly, this section puts more emphasis on these more vulnerable segments of the population.
15.4.5.1 Direct impacts of climate on the health of Arctic residents
Direct impacts (injury and death) are expected to result, in part, from exposure to temperature extremes and weather events. Increases in precipitation are expected to affect the frequency and magnitude of natural disasters such as debris flow, avalanches and rock falls (Koshida and Avis, 1998 [NotFound] ). Thunderstorms and high humidity are associated with short-term increases in respiratory and cardiovascular diseases ( Kovats et al., 2000 [NPR, ARC] ( Messner 2005 ) ) reported an increased incidence of non-fatal heart attacks with increased temperature during the positive phase of the Arctic Oscillation (AO) in Sweden, but related it to changes in behaviour that can cause an increase in the susceptibility of individuals to atherosclerotic diseases. Low temperatures and social stress have been related to cases of cardiomyopathy, a weakening of the heart muscle or change in heart muscle structure, identified in northern Russia ( Khasnullin et al., 2000 [NPR] ). Residents in some Arctic regions report respiratory stress associated with extreme warm summer days not previously experienced ( Furgal et al., 2002 [NPR, SRC] ).
The frequency of some injuries (e.g., frostbite, hypothermia) or accidents, and diseases (cardiovascular, respiratory, circulatory, musculoskeletal, skin) is increased by cold exposure ( (Hassi et al., 2005 ) ). An estimated 2,000 to 3,000 deaths/yr occur from cold-related diseases and injury in Finland during the cold season. This winter-related mortality is higher than the number of deaths associated with other common causes in the country throughout the year (e.g., 400/yr from traffic accidents, 100-200/yr from heat). The prevalence of respiratory diseases among children in the Russian North is 1.5 to 2 times higher than the national average. Evidence suggests that warming in Arctic regions during the winter months will reduce excess winter mortality, primarily through a reduction in cardiovascular and respiratory deaths ( (Nayha, 2005 ) ). Assuming that the standard of cold protection (including individual behavioural factors) does not deteriorate, a reduction in cold-related injuries is also likely ( (Nayha, 2005 ) ).
15.4.5.2 Indirect impacts of climate on the health of Arctic residents
Climate variability will have a series of more complex, indirect impacts on human-environment interactions in the Arctic ( Berner et al., 2005 [NPR, SRC] ). Local and traditional knowledge in nearly all regions records increasingly uncharacteristic environmental conditions and extremes not previously experienced (e.g., Krupnik and Jolly, 2002 [NPR] ). Evidence suggests that an increase in injuries among northern residents associated with ‘strange’ or changing environmental conditions, such as thinning and earlier break-up of sea ice, are related to trends in climate (e.g., Lafortune et al., 2004 [NPR, SRC] ).
Climate change in the Arctic during El Niño-Southern Oscillation (ENSO) events has been associated with illness in marine mammals, birds, fish and shellfish. A number of disease agents have been associated with these illnesses (e.g., botulism, Newcastle disease). It is likely that temperature changes arising from long-term climate change will be associated with an increased incidence of those diseases that can be transmitted to humans ( (Bradley et al., 2005 ) ). Many zoonotic diseases which currently exist in Arctic host species (e.g., tularemia in rabbits, rodents, muskrats and beaver, and rabies in foxes; Dietrich,). Many zoonotic diseases which currently exist in Arctic host species (e.g., tularemia in rabbits, rodents, muskrats and beaver, and rabies in foxes; Dietrich,) can spread via climate-controlled mechanisms (e.g., movement of animal populations). Similarly, the overwintering survival and distribution of many insect species that act as vectors of disease are positively impacted by warming temperatures and may mean that many diseases reappear, or new diseases appear, in Arctic regions ( (Parkinson and Butler, 2005 ) ). The examples of tick-borne encephalitis (brain infection) in Sweden ( (Lindgren and Gustafson, 2001 ) ), and Giardia spp. and Cryptosporidium spp. infection of ringed seals (Phoca hispida) and bowhead whales (Balaena mysticetus) in the Arctic Ocean are evidence of this potential ( (Hughes-Hanks et al., 2005 ) ).
Subsistence foods from the local environment provide Arctic residents with cultural and economic benefits and contribute a significant proportion of daily requirements of several vitamins and essential elements to the diet (e.g., ( Blanchet et al., 2000 ) ). Wild foods also comprise the greatest source of exposure to environmental contaminants. The uptake, transport and deposition behaviour of many of these chemicals is influenced by temperature, and therefore climate warming may indirectly influence human exposure ( Kraemer et al., 2005 [SRC] ). Through changes in accessibility and distribution of wildlife species, climate change in combination with other social, cultural, economic and political trends in Arctic communities, will be likely to influence the diet of circumpolar residents.
Transitions towards more market food items in Arctic indigenous diets to date have been associated with a rise in levels of cardiovascular diseases, diabetes, dental cavities and obesity ( Van Oostdam et al., 2003 [NPR] ). In many indigenous communities, these subsistence food systems are the basis of traditions, socio-economic and cultural well-being. Indigenous peoples maintain a strong connection to the environment through traditional resource-harvesting activities in a way that distinguishes them from non-indigenous communities, and this may indeed contribute to how specific peoples retain a fundamental identification to a particular area ( Gray, 1995 [NPR] ; Nuttall et al., 2005 [NPR, SRC] ).
While climate-related changes threaten aspects of food security for some subsistence systems, increased temperatures and decreased sea-ice cover represent increased transport opportunities and access to market food items. Shifts in animal population movements also mean potential introduction of new food species to northern regions. These combined effects on Arctic food security, in addition to increased opportunities for agricultural and pastoral activities with decreased severity of winter and lengthened summer growing seasons, make it difficult to predict how diets will change and impact health, even presupposing that we have a sufficient understanding of what local environments can provide and sustain. It is also clear that these impacts will be influenced not only by environmental change but also by economic, technological and political forces.
Through increased river and coastal flooding and erosion, increased drought, and degradation of permafrost, resulting in loss of reservoirs or sewage contamination, climate change is likely to threaten community and public health infrastructure, most seriously in low-lying coastal Arctic communities (e.g., Shishmaref, Alaska, USA; Tuktoyaktuk, Northwest Territories, Canada). Community water sources may be subject to salt-water intrusion and bacterial contamination. Quantities of water available for basic hygiene can become limited due to drought and damaged infrastructure. The incidence of disease caused by contact with human waste may increase when flooding and damaged infrastructure such as sewage lagoons, or inadequate hygiene, spreads sewage. However, treatment efficiencies in wastewater lagoons may also improve due to warmer water temperatures, delaying the need to expand natural wastewater treatment systems as local populations grow ( (Warren et al., 2005 ) ).
The combined socio-cultural, economic, political and environmental forces acting on and within Arctic communities today ( Chapin et al., 2005a [NPR, SRC] ) have significant implications for health and well-being ( (Curtis et al., 2005 ) ). Alterations in the physical environment threatening specific communities (e.g., through erosion and thawing permafrost) and leading to forced relocation of inhabitants, or shifts or declines in resources resulting in altered access to subsistence species (e.g., Inuit hunting of polar bear) can lead to rapid and long-term cultural change and loss of traditions. Such loss can, in turn, create psychological distress and anxiety among individuals ( (Hamilton et al., 2003; ) ( Curtis et al., 2005 ) ). However, across most of the Arctic, climate change is just one of many driving forces transforming communities. These forces arise from inside and outside the community, but combined are influencing the acculturation process by influencing ways of living, and loss of traditions that are positively related to social, cultural and psychological health ( (Berry, 1997 ) ).
The social, cultural and economic transitions that Arctic communities have seen over the last 50 years has influenced all aspects of health in the Arctic, and this influence is highly likely to continue in the future. Climate change is probably going to drive changes in communities by challenging individuals’ and communities’ relationships with their local environment, which has been the basis of Arctic peoples’ identity, culture, social and physical well-being (Einarsson et al., 2004 [NotFound] ; Berner et al., 2005 [NPR, SRC] ; Chapin et al., 2005a [NPR, SRC] ).
15.4.6 Coastal zone and small islands
15.4.6.1 Arctic coastal erosion
Coastal stability in polar regions is affected by factors common to all areas (exposure, relative sea-level change, climate and lithology), and by factors specific to the high latitudes (low temperatures, ground ice and sea ice). The most severe erosion problems affect infrastructure and culturally important sites in areas of rising sea level, where warming coincides with areas that are seasonally free of sea ice or where there is widespread ice-rich permafrost ( Forbes, 2005 [NPR, SRC] ). Ice-rich permafrost is widespread in the western Canadian Arctic, northern Alaska and along much of the Russian Arctic coast (e.g., ( Smith, 2002; ) Nikiforov et al., 2003 [NPR] ). Wave erosion and high summer air temperatures promote rapid shoreline retreat, in some cases contributing a significant proportion of regional sediment and organic carbon inputs to the marine environment ( Aré, 1999 [NPR] ; Rachold et al., 2000 [PoC, JoC] ). Communities located on resistant bedrock or where glacio-isostatic rebound is occurring are less vulnerable to erosion.
Coastal instability may be further magnified by poorly adapted development. For example, in places such as Varandey (Russian Federation) industrial activity has promoted erosion, leading to the destruction of housing estates and industrial facilities ( (Ogorodov, 2003 ) ). Interacting human and natural effects may also increase the sensitivity to coastal erosion. For example, in Shishmaref (Alaska, USA) and Tuktoyaktuk (Northwest Territories, Canada), the combined effects of reduced sea ice, thawing permafrost, storm surges and waves have led to significant loss of property, and this has led to relocation or abandonment of homes and other facilities ( Instanes et al., 2005 [NPR, SRC] ). Despite a cultural aversion to moving from traditional sites, these changes may ultimately force relocation. Although clear evidence for accelerated erosion is sparse, there has been a documented increase in erosion rates between 1954 - 1970 and 1970 - 2000 for coastal terrain with very high ground-ice content at Herschel Island, Canada ( (Lantuit and Pollard, 2003 ) ). A modelling exercise ( Rasumov, 2001 [MoS] ) suggested that erosion rates in the eastern Siberian Arctic could increase by 3-5 m/yr with a 3°C increase in mean summer air temperature. Furthermore, the projected reduction of sea ice would also contribute to increased erosion, as has been observed at Nelson Lagoon in Alaska ( Instanes et al., 2005 [NPR, SRC] ).
15.4.6.2 Sub-Antarctic islands
Several sub-Antarctic islands have undergone substantial recent climate change, the impacts of which have been significant and have included profound physical and biological changes (specific examples can be found in Chapter 11 , Sections 11.2.1 and 11.4.2 , Table 11.1 ; Chapter 16 , Section 16.4.4 ).
15.5 Adaptation: practices, options and constraints
Circum-Arctic nations are responsible for a significant fraction of global CO2 emissions and the Arctic is an important source of fossil fuels. Although some residents and less developed regions may contribute only a very small proportion of these nations’ emissions, there is a need to consider both mitigation and adaptation in polar regions in the light of trends in resource development and modernisation taking place in these areas. The burden faced by Arctic residents is magnified by the observed and projected amplification of climate change in the Arctic and the potential for dramatic environmental impacts. As with other vulnerable regions of the world, human adaptation is critical, particularly for those living in closest relationship with the local environment.
Historically, cultural adaptations and the ability of Arctic indigenous peoples to utilise their local resources have been associated with, or affected by, seasonal variation and changing ecological conditions. One of the hallmarks of successful adaptation has been flexibility in technology and social organisation, and the knowledge and ability to cope with climate change and circumvent some of its negative impacts. Indigenous groups have developed resilience through sharing resources in kinship networks that link hunters with office workers, and even in the cash sector of the economy. Many people work flexibly, changing jobs frequently and having several part-time jobs ( Chapin et al., 2006 [JoC, SRC] ). Historically, responses to major climatic and environmental changes included an altering of group size or moving to appropriate new locations, flexibility with regard to seasonal cycles and harvesting, and the establishment of sharing mechanisms and networks for support ( Krupnik, 1993 [NPR] ; Freeman, 1996 [NPR] ). Many of these strategies, with the exception of group mobility, are still employed in various forms today (e.g., ( Berkes and Jolly, 2001; ) Nickels et al., 2002 [NPR, SRC] ; McCarthy et al., 2005 [NPR] ) yet, in the future, such responses may be constrained by social, cultural, economic and political forces acting on communities externally and from within.
Detailed local knowledge and the social institutions in which it exists are critical foundations of understanding interactions between people and their environment and therefore vital to community adaptability (see Section 15.6.1 ). Yet the generation of this knowledge requires active engagement with the environment and, as the nature of this interaction changes (e.g., amount and frequency of time spent on land or engaged in subsistence activities), so does the information it provides. Changes in local environments further challenge this knowledge and can increase human vulnerability to climatic and social change.
Greater uncertainty and threats to food security stress the need for resilient and flexible resource procurement systems. Resilience and adaptability depend on ecosystem diversity as well as the institutional rules that govern social and economic systems ( (Adger, 2000 ) ). Innovative co-management of both renewable and non-renewable resources could support adaptive abilities via flexible management regimes while providing opportunities to enhance local economic benefits and ecological and societal resilience ( Chapin et al., 2004 [SRC] ).
Opportunities for adaptation exist within some changes already taking place. The arrival of new species (e.g., Babaluk et al., 2000 [SRC] ; Huntington et al., 2005 [NPR] ) and an increase in growing seasons and opportunities for high-latitude agriculture provide opportunities to enhance resilience in local food systems. Increased eco-tourism may increase incentives for protection of environmental areas. Taking advantage of these potentially positive impacts will, however, require institutional flexibility and forms of economic support.
Given the interconnected nature of Arctic ecosystems and human populations, strategies are required that take a broad approach to support adaptation among a range of sectors. For example, policies that allow local people to practice subsistence activities within protected areas contribute to both biodiversity and cultural integrity ( Chapin et al., 2005a [NPR, SRC] ). The creation and protection of critical areas such as parks, with flexible boundaries to compensate for changing climatic conditions, enhances conservation of wildlife and services provided by this land for human use (e.g., tourism and recreation) ( Chapin et al., 2005a [NPR, SRC] ).
Although Arctic communities in many regions show great resilience and ability to adapt, some responses have been compromised by socio-political change. The political, cultural and economic diversity that exists among Arctic regions today impacts how communities are affected by, and respond to, environmental change. Such diversity also means that particular experiences of climate variability, impacts and responses may not be universal. Currently, little is known about how communities and individuals, indigenous or non-indigenous, differ in the way risks are perceived, or how they might adapt aspects of their lives (e.g., harvesting strategies) in response to negative change. The effectiveness of local adaptive strategies is uneven across the Arctic and there are large gaps in knowledge about why some communities do well, while others are more vulnerable to drivers of change, even when they share similar resources and ecological settings. Ultimately, an understanding of adaptation can only derive from a better understanding of social and economic vulnerability among all Arctic residents ( Handmer et al., 1999 [ARC] ).
15.6 Case studies
15.6.1 Case study: traditional knowledge for adaptation
Among Arctic peoples, the selection pressures for the evolution of an effective knowledge base have been exceptionally strong, driven by the need to survive off highly variable natural resources in the remote, harsh Arctic environment. In response, they have developed a strong knowledge base concerning weather, snow and ice conditions as they relate to hunting and travel, and natural resource availability ( Krupnik and Jolly, 2002 [NPR] ). These systems of knowledge, belief and practice have been developed through experience and culturally transmitted among members and across generations ( (Huntington, 1998; ) Berkes, 1999 [NPR] ). This Arctic indigenous knowledge offers detailed information that adds to conventional science and environmental observations, as well as to a holistic understanding of environment, natural resources and culture ( Huntington et al., 2004 [SRC] ). There is an increasing awareness of the value of Arctic indigenous knowledge and a growing collaborative effort to document it. In addition, this knowledge is an invaluable basis for developing adaptation and natural resource management strategies in response to environmental and other forms of change. Finally, local knowledge is essential for understanding the effects of climate change on indigenous communities ( (Riedlinger and Berkes, 2001; ) Krupnik and Jolly, 2002 [NPR] ) and how, for example, some communities have absorbed change through flexibility in traditional hunting, fishing and gathering practices.
The generation and application of this knowledge is evidenced in the ability of Inuit hunters to navigate new travel and hunting routes despite decreasing ice stability and safety (e.g., Lafortune et al., 2004 [NPR, SRC] ); in the ability of many indigenous groups to locate and hunt species such as geese and caribou that have shifted their migration times and routes and to begin to locate and hunt alternative species moving into the region (e.g., Krupnik and Jolly, 2002 [NPR] ; Nickels et al., 2002 [NPR, SRC] ; Huntington et al., 2005 [NPR] ); the ability to detect safe sea ice and weather conditions in an environment with increasingly uncharacteristic weather ( (George et al., 2004 ) ); or the knowledge and skills required to hunt marine species in open water later in the year under different sea ice conditions (Community of Arctic Bay, 2005 ).
Although Arctic peoples show great resilience and adaptability, some traditional responses to environmental change have already been compromised by recent socio-political changes. Their ability to cope with substantial climatic change in future, without a fundamental threat to their cultures and lifestyles, cannot be considered as unlimited. The generation and application of traditional knowledge requires active engagement with the environment, close social networks in communities, and respect for and recognition of the value of this form of knowledge and understanding. Current social, economic and cultural trends, in some communities and predominantly among younger generations, towards a more western lifestyle has the potential to erode the cycle of traditional knowledge generation and transfer, and hence its contribution to adaptive capacity.
15.6.2 Case study: Arctic megadeltas
Numerous river deltas are located along the Arctic coast and the rivers that flow to it. Of particular importance are the megadeltas of the Lena (44,000 km2) and Mackenzie (9,000 km2) rivers, which are fed by the largest Arctic rivers of Eurasia and North America, respectively. In contrast to non-polar megadeltas, the physical development and ecosystem health of these systems are strongly controlled by cryospheric processes and are thus highly susceptible to the effects of climate change.
Currently, advance/retreat of Arctic marine deltas is highly dependent on the protection afforded by near-shore and land-fast sea ice ( Solomon, 2005 [PoC, JoC] ; Walsh et al., 2005 [NPR, PoC, SRC] ). The loss of such protection with warming will lead to increased erosion by waves and storm surges. The problems will be exacerbated by rising sea levels, greater wind fetch produced by shrinking sea-ice coverage, and potentially by increasing storm frequency. Similarly, thawing of the permafrost and ground-ice that currently consolidates deltaic material will induce hydrodynamic erosion on the delta front and along riverbanks. Thawing of permafrost on the delta plain itself will lead to similar changes; for example, the initial development of more ponded water, as thermokarst activity increases, will eventually be followed by drainage as surface and groundwater systems become linked. Climate warming may have already caused the loss of wetland area as lakes expanded on the Yukon River delta in the late 20th century ( Coleman and Huh, 2004 [NPR] ). Thaw subsidence may also affect the magnitude and frequency of delta flooding from spring flows and storm surges ( (Kokelj and Burn, 2005 ) ).
The current water budget and sediment-nutrient supply for the multitude of lakes and ponds that populate much of the tundra plains of Arctic deltas depends strongly on the supply of floodwaters produced by river-ice jams during the spring freshet. Studies of future climate conditions on a major river delta of the Mackenzie River watershed (Peace-Athabasca Delta) indicate that a combination of thinner river ice and reduced spring runoff will lead to decreased ice-jam flooding ( Beltaos et al., 2006 [SRC] ). This change combined with greater summer evaporation, due to warmer temperatures, will cause a decline in delta-pond water levels ( (Marsh and Lesack, 1996 ) ). For many Arctic regions, summer evaporation already exceeds precipitation and therefore the loss of ice-jam flooding could lead to a drying of delta ponds and a loss of sediment and nutrients known to be critical to their ecosystem health ( (Lesack et al., 1998; ) ( Marsh et al., 1999 ) ). A successful adaptation strategy that has already been used to counteract the effects of drying of delta ponds involves managing water release from reservoirs to increase the probability of ice-jam formation and related flooding ( Prowse et al., 2002 [Ambiguous] ).
15.6.3 Case study: Antarctic Peninsula – rapid warming in a pristine environment
The Antarctic Peninsula is a rugged mountain chain generally more than 2,000 m high, differing from most of Antarctica by having a summer melting season. Summer melt produces many isolated snow-free areas, which are habitats for simple biological communities of primitive plants, microbes and invertebrates, and breeding grounds for marine mammals and birds. The Antarctic Peninsula has experienced dramatic warming at rates several times the global mean ( Vaughan et al., 2003 [JoC, SRC] ; Trenberth et al., 2007 [NPR, PoC, 2007] ). Since the TAR, substantial progress has been made in understanding the causes and profound impacts of this warming.
Since records began, 50 years ago, mean annual temperatures on the Antarctic Peninsula have risen rapidly; >2.5°C at Vernadsky (formerly Faraday) Station ( Turner et al., 2005 [JoC] ). On the west coast, warming has been much slower in summer and spring than in winter or autumn, but has been sufficient to raise the number of positive-degree-days by 74% ( Vaughan et al., 2003 [JoC, SRC] ), and the resulting increase in melt has caused dramatic impacts on the Antarctic Peninsula environment, and its ecology.
Around 14,000 km2 of ice have been lost from ten floating ice shelves ( King, 2003 [NPR] ), 87% of glacier termini have retreated ( Cook et al., 2005 [JoC, SRC] ), and seasonal snow cover has decreased ( (Fox and Cooper, 1998 ) ). The loss of seasonal snow and floating ice do not have a direct impact on global sea level, but acceleration of inland glaciers due to the loss of ice shelves ( De Angelis and Skvarca, 2003 [JoC] ; Scambos et al., 2004 [JoC] ; Rignot et al., 2005 [JoC, SRC] ) and increased run-off of melt water ( Vaughan, 2006 [SRC] ) will cause an increase in this contribution. If summer warming continues, these effects will grow.
Marine sediment cores show that ice shelves probably have not reached a similar minimum for at least 10,000 years ( Domack et al., 2005 [JoC] ), and certainly not for 1,000 years ( (Pudsey and Evans, 2001; ) Domack et al., 2003 [NPR] ). This suggests that the retreat is not simply due to cyclic variations in local climate, and that recent warming is unique in the past 10,000 years ( Turner et al., 2007 [JoC, SRC, 2007] ). The processes leading to warming are unclear, but appear to be correlated with atmospheric circulation ( van den Broeke and van Lipzig, 2003 [NPR] ) and particularly with changes in the Southern Annular Mode caused by anthropogenic influence ( Marshall et al., 2004 [JoC] ; Marshall et al., 2006 [JoC] ). The winter warming on the west coast also appears to be related to persistent retreat of sea ice (see Figure 15.2 ( Parkinson, 2002 ) ) and warming in the Bellingshausen Sea ( Meredith and King, 2005 [JoC] ). The spring depletion of ozone over Antarctica (the Antarctic Ozone Hole) has also been implicated in driving circulation change ( Thompson and Solomon, 2002 [PoC, JoC] ), but this has been disputed ( Marshall et al., 2004 [JoC] ). Current general circulation models (GCMs) do not, however, simulate this observed warming over the past 50 years ( King, 2003 [NPR] ) and we cannot predict with confidence whether rapid warming will continue in future.
If warming does continue (especially in the summer) there will be significant impacts; retreat of coastal ice and loss of snow cover would result in newly exposed rock and permafrost – providing new habitats for colonisation by expanding and invading flora and fauna. However, the direct impacts of climate change on the flora and fauna are difficult to predict, since these ecosystems are subject to multiple stressors. For example, increased damage by ultraviolet exposure, because of reduced ozone levels and summer desiccation, may oppose the direct responses to warming ( (Convey et al., 2002 ) ). In addition, there is a growing threat of alien species invasion, as climatic barriers to their establishment are eroded by climate amelioration, and increasing human activity increases the opportunity for introduction. Such invasions have already occurred on many sub-Antarctic islands, with detrimental consequences for native species ( (Frenot et al., 2005 ) ). Furthermore, slow reproduction rates during rapid climate change may limit the possible relocation of native species.
There have been trends in all trophic levels in the marine ecosystems west of the Antarctic Peninsula. These have been driven by reduced sea-ice extent and duration. Changes in primary production may also have been affected by increases in the supply of glacial melt ( Smith et al., 2003 [NPR] ). Similarly, reduced sea-ice cover was the likely cause of the dramatic change in the balance between krill and salps, the main grazers of phytoplankton ( Atkinson et al., 2004 [JoC] ). The loss of krill will probably have impacts on higher predators (albatrosses, seals, whales and penguins: populations of the latter are already changing; Smith et al., 2003 [NPR] ), but could have more far-reaching impacts, perhaps even affecting CO2 sequestration in parts of the Southern Ocean ( (Walsh et al., 2001 ) ).
The global significance of the Antarctic Peninsula warming is difficult to encapsulate, but the main concern is for the loss of a unique landscape and biota. The rate of warming on the Antarctic Peninsula is among the highest seen anywhere on Earth in recent times, and is a dramatic reminder of how subtle climate-dynamic processes can drive regional climate change, and the complexity of its impacts in an environment where human influence is at a minimum.
15.7 Conclusions: implications for sustainable development
15.7.1 Economic activity, infrastructure and sustainability in the Arctic
The thawing of ice-rich permafrost creates potential for subsidence and damage to infrastructure, including oil and gas extraction and transportation facilities ( Hayley, 2004 [NPR] ), and climate warming will exacerbate existing subsidence problems ( Instanes et al., 2005 [NPR, SRC] ). These risks have been assessed using a ‘permafrost hazard’ index (e.g., Nelson et al., 2001 [JoC, SRC] ; Anisimov and Belolutskaia, 2004 [MoS, SRC] ; Anisimov and Lavrov, 2004 [SRC] ; Smith and Burgess, 2004 [NPR] ), which, when coupled with climate projections, suggests that a discontinuous high-risk zone (containing population centres, pipelines and extraction facilities) will develop around the Arctic Ocean by the mid-21st century ( Nelson et al., 2001 [JoC, SRC] ). Similarly, a zone of medium risk contains larger population centres (Yakutsk, Noril?sk, Vorkuta) and much of the Trans-Siberian and Baikal-Amur railways. However, distinguishing between the broad effects of climate change on permafrost and more localised human-induced changes remains a significant challenge ( (Tutubalina and Rees, 2001; ) Nelson, 2003 [JoC, SRC] ). Although several recent scientific and media reports have linked widespread damage to infrastructure with climate change (e.g., Smith et al., 2001 [NPR] ; Couture et al., 2003 [NPR, PoC] ), the effect of heated buildings on underlying ice-rich permafrost can easily be mistaken for a climate-change impact. Similarly, urban heat-island effects occur in northern settlements (e.g., Hinkel et al., 2003 [JoC, SRC] ) and may be a factor in local degradation of permafrost.
The cost of rehabilitating community infrastructure damaged by thawing of permafrost could be significant ( (Couture et al., 2000, ) 2001; Chartrand et al., 2002 [NPR] ). Even buildings designed specifically for permafrost environments may be subject to severe damage if design criteria are exceeded ( Khrustalev, 2000 [NPR] ). The impervious nature of ice-rich permafrost has been relied on as a design element in landfill and contaminant-holding facilities ( (Snape et al., 2003 ) ), and thawing such areas could result in severe contamination of hydrological resources and large clean-up costs, even for relatively small spills ( (Roura, 2004 ) ). Rates of coastal erosion in areas of ice-rich permafrost are among the highest anywhere and could be increased by rising sea levels ( Brown et al., 2003 [NPR] ). Relocation of threatened settlements would incur very large expenses. It has been estimated that relocating the village of Kivalina, Alaska, to a nearby site would cost US$54 million (U.S. Arctic Research Commission Permafrost Task Force, 2003 [NotFound] ). However, some fraction of the costs will be offset by economic benefits to northern communities. For example, there will be savings on heating costs: modelling has predicted a 15% decline in the demand for heating energy in the populated parts of the Arctic and sub-Arctic and up to 1 month decrease in the duration of the period when heating is needed ( Anisimov, 1999 [SRC] ).
Lakes and river ice have historically provided major winter transportation routes and connections to smaller settlements. Reductions in ice thickness will reduce the load-bearing capacity, and shortening of the ice season will shorten periods of access. Adaptation in the initial stages of climate change will be through modified construction techniques and transport vehicles and schedules, but longer-term strategies will require new transportation methods and routes. Where an open-water network is viable, it will be sensible to increase reliance on water transport. In land-locked locations, the construction of all-weather roads may be the only viable option, with implications for significantly increased costs (e.g., Lonergan et al., 1993 [JoC] ; Dore and Burton, 2001 [NPR, MoS, ARC] ). Similar issues will impact the use of the sea-ice roads primarily used to access offshore facilities.
Loss of summer sea ice will bring an increasingly navigable Northwest Passage, and the Northern Sea Route will create new opportunities for cruise shipping. Projections suggest that by 2050, the Northern Sea Route will have 125 days/yr with less than 75% sea-ice cover, which represents favourable conditions for navigation by ice-strengthened cargo ships ( Instanes et al., 2005 [NPR, SRC] ). Increased marine navigation and longer summers will improve conditions for tourism and travel associated with research ( Instanes et al., 2005 [NPR, SRC] ), and this effect is already being reported in the North American Arctic ( Eagles, 2004 [NPR] ).
Even without climate change, the complexity of producing a viable plan for sustainable development of the Arctic would be daunting; but the added uncertainty of climate change, and its likely amplification in the Arctic, make this task enormous. The impacts on infrastructure discussed above, together with the probable lengthening of growing seasons and increasing agricultural effort, opening of new sea routes, changing fish stocks, and ecosystem changes will provide many new opportunities for the development of Arctic economies. However it will also place limits on how much development is actually sustainable. There does, however, now appear to be an increasing understanding, among governments and residents, that environmental protection and sustainable development are two sides of the same coin ( Nuttall, 2000a [NPR, SRC] ), and a forum for circum-Arctic co-operation exists in the Arctic Council. This involves eight nations and six indigenous peoples’ organisations and embraces the concept of sustainable development in its mandate. The Arctic Council, in partnership with the International Arctic Science Committee, is responsible for the recent Arctic Climate Impact Assessment ( ACIA, 2005 [NPR] ), which has substantially improved the understanding of the impacts of climate change in the Arctic, is a benchmark for regional impact assessments, and may become the basis for a sustainable management plan for the Arctic.
15.7.2 Economic activity and sustainability in the Antarctic
Fishing and tourism are the only significant economic activities in the Antarctic at present. Over 27,000 tourists visited Antarctica in the 2005 /06 summer and the industry is growing rapidly ( IAATO, 2006 [NPR] ). The multiple stresses of climate change and increasing human activity on the Antarctic Peninsula represent a clear vulnerability (see Section 15.6.3 ), and have necessitated the implementation of stringent clothing decontamination guidelines for tourist landings on the Antarctic Peninsula ( IAATO, 2005 [NPR] ).
Fishing is, however, the only large-scale exploitation of resources in Antarctica, and since 1982 Antarctic fisheries have been regulated by the Convention on the Conservation of Antarctic Marine Living Resources (CCAMLR), which takes climate change into account in determining allowable catches. However, before the CCAMLR came into force, heavy fishing around South Georgia led to a major decline in some stocks, which have not yet fully recovered. The illegal, unregulated and unreported fishing of the Patagonian toothfish (Dissostichus eleginoides) is of concern because it could act alongside climate change to undermine sustainable management of stocks ( (Bialek, 2003 ) ). Furthermore, those fishing illegally often use techniques that cause the death of by-catch species; for example, albatross and petrels, which are now under threat ( Tuck et al., 2001 [MoS] ).
15.8 Key uncertainties and research priorities
Significant advances in our understanding of polar systems have been made since the TAR ( Anisimov et al., 2001 [Ambiguous] ) and the Arctic Climate Impacts Assessment reports ( ACIA, 2005 [NPR] ). Many climate-induced changes that were anticipated in the TAR have now been documented. This validation, together with improved models, new data and increasing use of indigenous and local knowledge, has increased our confidence in projecting future changes in the polar regions, although substantial uncertainties remain, and the remote and harsh environments of the polar regions constrain data collection and mean that observational networks are sparse and mostly only recently established. The difficulty in understanding climate-change effects in polar regions is further exacerbated by the complexity within and among polar systems, their feedbacks and sensitivity, and their potential to switch into different states of (dis)equilibrium.
Significant research since the TAR has focused on the impact of climate change on Arctic indigenous populations, and accordingly, this chapter has placed an emphasis on these segments of the population. However, the impacts on the wider population need also be considered, and in particular, the economic impacts, which are difficult to address at present due to the dearth of information.
To address the key uncertainties, particular approaches will be required ( Table 15.1 ). For the Arctic, detailed recommendations for future research have been drafted by the international scientific community (ICARP II, 2006 ), and a burst of co-ordinated research at both poles is anticipated during the International Polar Year, 2007 - 2009 .
Table 15.1. Key uncertainties and related scientific recommendations/approaches.
| Uncertainty | Recommendation and approach |
|---|---|
| Detection and projection of changes in terrestrial, freshwater and marine Arctic and Antarctic biodiversity and implications for resource use and climatic feedbacks | Further development of integrated monitoring networks and manipulation experiments; improved collation of long-term data sets; increased use of traditional knowledge and development of appropriate models |
| Current and future regional carbon balances over Arctic landscapes and polar oceans, and their potential to drive global climate change | Expansion of observational and monitoring networks and modelling strategies |
| Impacts of multiple drivers (e.g., increasing human activities and ocean acidity) to modify or even magnify the effects of climate change at both poles | Development of integrated bio-geophysical and socio-economic studies |
| Fine-scaled spatial and temporal variability of climate change and its impacts in regions of the Arctic and Antarctic | Improved downscaling of climate predictions, and increased effort to identify and focus on impact ‘hotspots’ |
| The combined role of Arctic freshwater discharge, formation/melt of sea ice and melt of glaciers/ice sheets in the Arctic and Antarctic on global marine processes including the thermohaline circulation | Integration of hydrologic and cryospheric monitoring and research activities focusing on freshwater production and responses of marine systems |
| The consequences of diversity and complexity in Arctic human health, socio-economic, cultural and political conditions; interactions between scales in these systems and the implications for adaptive capacity | Development of standardised baseline human system data for circumpolar regions; integrated multidisciplinary studies; conduct of sector-specific, regionally specific human vulnerability studies |
| Model projections of Antarctic and Arctic systems that include thresholds, extreme events, step-changes and non-linear interactions, particularly those associated with phase-changes produced by shrinking cryospheric components and those associated with disturbance to ecosystems | Appropriate interrogation of existing long-term data sets to focus on non-linearities; development of models that span scientific disciplines and reliably predict non-linearities and feedback processes |
| The adaptive capacity of natural and human systems to cope with critical rates of change and thresholds/tipping points | Integration of existing human and biological climate-impact studies to identify and model biological adaptive capacities and formulate human adaptation strategies |
References
Aanes, R., B.-E. Sæther and N.A. Øritsland, 2000: Fluctuations of an introduced population of Svalbard reindeer: the effects of density dependence and climatic variation. Ecography, 23, 437-443. Clean
Abdalati, W. and K. Steffen, 2001: Greenland ice sheet melt extent: 1979-1999. J. Geophys. Res., 106, 33983-33988. [JoC]
ACIA, 2005: Impacts of a Warming Arctic: Arctic Climate Impacts Assessment. Cambridge University Press, Cambridge, 1042 pp. [NPR]
Ackley, S., P. Wadhams, J.C. Comiso and A.P. Worby, 2003: Decadal decrease of Antarctic sea ice extent inferred from whaling records revisited on the basis of historical and modern sea ice records. Polar Res., 22, 19-25. Clean
Adger, W.N., 2000: Social and ecological resilience: are they related? Prog. Hum. Geog., 24, 347-364. Clean
Agnarsson, S. and R. Arnason, 2003: The role of the fishing industry in the Icelandic economy: an historical examination. Working Paper W03:07. Institute of Economic Studies, University of Iceland, Reykjavik, 24 pp. [NPR]
Ainley, D.G., E.D. Clarke, K. Arrigo, W.R. Fraser, A. Kato, K.J. Barton and P.R. Wilson, 2005: Decadal-scale changes in the climate and biota of the Pacific sector of the Southern Ocean, 1950s to the 1990s. Antarct. Sci., 17, 171-182. Clean
Albon, S.D., A. Stien, R.J. Irvine, R. Langvatn, E. Ropstad and O. Halvorsen, 2002: The role of parasites in the dynamics of a reindeer population. P. Roy. Soc. Lond. B, 269, 1625-1632. Clean
Anisimov, O., 1999: Impact of changing climate on building heating and air conditioning. Meteorol. Hydrol., 6, 10-17. [SRC]
Anisimov, O.A. and M.A. Belolutskaia, 2001: Predicting agroclimatic parameters using geographical information system. Meteorol. Hydrol., 9, 89-98. [MoS, SRC]
Anisimov, O.A. and M.A. Belolutskaia, 2004: Predictive modelling of climate change impacts on permafrost: effects of vegetation. Meteorol. Hydrol., 11, 73-81. [MoS, SRC]
Anisimov, O.A. and C.A. Lavrov, 2004: Global warming and permafrost degradation: risk assessment for the infrastructure of the oil and gas industry. Technologies of Oil and Gas Industry, 3, 78-83. [SRC]
Anisimov, O., B.B. Fitzharris, J.O. Hagen, B. Jefferies, H. Marchant, F. Nelson, T. Prowse and D. Vaughan, 2001: Polar regions (Arctic and Antarctic). Climate Change 2001: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change, J.J. McCarthy, O.F. Canziani, N.A. Leary, D.J. Dokken and K.S. White, Eds., Cambridge University Press, Cambridge, 801-841. [NPR, SRC]
Anisimov, O.A., S.A. Lavrov and S.A. Reneva, 2005a: Emission of methane from the Russian frozen wetlands under the conditions of the changing climate. Problems of Ecological Modeling and Monitoring of Ecosystems, Yu. Izrael, Ed., Hydrometeoizdat, St. Petersburg, 124-142. [NPR, MoS, SRC]
Anisimov, O.A., S.A. Lavrov and S.A. Reneva, 2005b: Modelling the emission of greenhouse gases from the Arctic wetlands under the conditions of the global warming. Climatic and Environmental Changes, G.V. Menzhulin, Ed., Hydrometeoizdat, St. Petersburg, 21-39. [NPR, SRC]
Arctic Monitoring and Assessment Programme, 2003: AMAP Assessment 2002: Human Health in the Arctic. Arctic Monitoring and Assessment Programme, Oslo, 137 pp. [NPR]
Aré, F.E., 1999: The role of coastal retreat for sedimentation in the Laptev Sea. Land–Ocean Systems in the Siberian Arctic: Dynamics and History, H. Kassens, H.A Bauch, I.A. Dmitrenko, H. Eicken, H.-W. Hubberten, M. Melles, J. Thiede and L.A. Timikhov, Eds., Springer, Berlin, 287-299. [NPR]
Arora, V.K. and G.J. Boer, 2001: Effects of simulated climate change on the hydrology of major river basins. J. Geophys. Res., 106, 3335-3348. [JoC, MoS]
Atkinson, A., V. Siegel, E. Pakhomov and P. Rothery, 2004: Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature, 432, 100-103. [JoC]
Atkinson, D.E., 2005: Environmental forcing of the circum-Polar coastal regime. Geo-Marine Lett., 25, 98-109. [JoC, MoS]
Aurela, M., T. Laurila and J.-P. Tuovinen, 2002: Annual CO2 balance of a subarctic fen in northern Europe: importance of the wintertime efflux. J. Geophys. Res., 107, 4607, doi:10.1029/2002JD002055. [JoC]
Babaluk, J.A., J.D. Reist, J.D. Johnson and L. Johnson, 2000: First records of sockeye (Oncorhynchus nerka) and pink salmon (O. gorbuscha) from Banks Island and other records of pacific salmon in Northwest Territories, Canada. Arctic, 53, 161-164. [SRC]
Beerling, D.J., A.C. Terry, P.L. Mitchell, D. Gwynn-Jones, J.A. Lee and T.V. Callaghan, 2001: Time to chill: effects of simulated global change on leaf ice nucleation temperatures of sub-Arctic vegetation. Am. J. Bot., 88, 628-633. [MoS, SRC]
Beilman, D.W. and S.D. Robinson, 2003: Peatland permafrost thaw and landform type along a climatic gradient. Proc., 8th International Conference on Permafrost, M. Phillips, S.M. Springman and L.U. Arenson, Eds., A.A. Balkema, Lisse, 61-66. [NPR]
Beilman, D.W., D.H. Vitt and L.A. Halsey, 2001: Localized permafrost peatlands in western Canada: definition, distributions, and degradation. Arct. Antarct. Alp. Res., 33, 70-77. Clean
Beltaos, S., T. Prowse, B. Bonsal, R. MacKay, L. Romolo, A. Pietroniro and B. Toth, 2006: Climatic effects on ice-jam flooding of the Peace–Athabasca Delta. Hydrol. Process., 20, 4013-4050. [SRC]
Berezovskaya, S., D.Q. Yang and L. Hinzman, 2005: Long-term annual water balance analysis of the Lena River. Global Planet. Change, 48, 84-95. Clean
Bergstrom, D.M. and S.L. Chown, 1999: Life at the front: history, ecology and change on southern ocean islands. Trends Ecol. Evol., 14, 472-477. Clean
Berkes, F., 1999: Sacred Ecology: Traditional Ecological Knowledge and Resource Management. Taylor and Francis, London, 232 pp. [NPR]
Berkes, F., 2002: Epilogue: making sense of arctic environmental change? The Earth is Faster Now: Indigenous Observations of Arctic Environmental Change, I. Krupnik and D. Jolly, Eds., Arctic Research Consortium, Fairbanks, 335-349. [NPR]
Berkes, F. and D. Jolly, 2001: Adapting to climate change: social-ecological resilience in a Canadian Western Arctic Community. Conserv. Ecol., 5, 18. Clean
Berner, J., C. Furgal, P. Bjerregaard, M. Bradley, T. Curtis, E. De Fabo, J. Hassi, W. Keatinge, S. Kvernmo, S. Nayha, H. Rintamaki and J. Warren, 2005: Human health. Arctic Climate Impact Assessment, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 863-906. [NPR, SRC]
Berry, J.W., 1997: Immigration, acculturation and adaptation. Appl. Psychol., 46, 5-34. Clean
Betts, A.K. and J.H. Ball, 1997: Albedo over the boreal forest. J. Geophys. Res., 102, 28901-28909. [JoC]
Betts, R.A., 2000: Offset of the potential carbon sink from boreal forestation by decreases in surface albedo. Nature, 408, 187-190. [JoC, ARC]
Bialek, D., 2003: Sink or swim: measures under international law for the conservation of the Patagonian toothfish in the Southern Ocean. Ocean Dev. Int. Law, 34, 105-137. Clean
Bindoff, N., J. Willebrand, V. Artale, A. Cazenave, J. Gregory, S. Gulev, K. Hanawa, C. Le Quere, S. Levitus, Y. Nojiri, C. Shum, L. Talley and U. Alakkat, 2007: Observations: oceanic climate change and sea level. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor and H.L. Miller, Eds., Cambridge University Press, Cambridge, 385-432. [NPR, 2007]
Blanchet, C., E. Dewailly, P. Ayotte, S. Bruneau, O. Receveur and B. Holub, 2000: Contribution of selected traditional and market foods to the diet of Nunavik Inuit women. Can. J. Diet. Pract. Res., 61, 1-9. Clean
Bochkov, Y.A., 1982: Water temperature in the 0-200 m layer in the Kola Meridian Section in the Barents Sea, 1900-1981 (in Russian). Sb. Nauch. Trud. PINRO, 46, 113-122. Clean
Bogaart, P.W. and R.T. van Balen, 2000: Numerical modeling of the response of alluvial rivers to Quaternary climatic change. Global Planet. Change, 27, 124-141. [MoS]
Bogoyavlenskiy, D. and A. Siggner, 2004: Arctic demography. Arctic Human Development Report (AHDR), N. Einarsson, J.N. Larsen, A. Nilsson and O.R. Young, Eds., Steffanson Arctic Institute, Akureyri, 27-41. [NPR]
Bonan, G.B., D. Pollard and S.L. Thompson, 1992: Effects of boreal forest vegetation on global climate. Nature, 359, 716-718. [JoC]
Both, C., R.G. Bijlsma and M.E. Visser, 2005: Climate effects on timing of spring migration and breeding in a long-distance migrant, the pied flycatcher, Ficedula hypoleuca. J. Avian Biol., 36, 368-373. [NPR]
Bradley, M.J., S.J. Kutz, E. Jenkins and T.M. O’Hara, 2005: The potential impact of climate change on infectious diseases of Arctic fauna. International Journal for Circumpolar Health, 64, 468-477. Clean
Brown, J., M.T. Jorgenson, O.P. Smith and W. Lee, 2003: Long-term rates of coastal erosion and carbon input, Elson Lagoon, Barrow, Alaska. Proc., 8th International Conference on Permafrost, M. Phillips, S.M. Springman and L.U. Arenson, Eds., A.A. Balkema, Lisse, 101-106. [NPR]
Buch, E., S.A. Horsted and H. Hovgaard, 1994: Fluctuations in the occurrence of cod in Greenland waters and their possible causes. ICES Mar. Sci. Symp., 198, 158-174. Clean
Callaghan, T.V., M. Johansson, O.W. Heal, N.R. Sælthun, L.J. Barkved, N. Bayfield, O. Brandt, R. Brooker and Co-authors, 2004: Environmental changes in the North Atlantic region: SCANNET as a collaborative approach for documenting, understanding and predicting changes. Ambio, 13, S39-S50. [MoS, SRC]
Callaghan, T.V., L.O. Björn, Y.I. Chernov, F.S. Chapin III, T.R. Christensen, B. Huntley, R. Ims, M. Johansson, D. Jolly, N.V. Matveyeva, N. Panikov, W.C. Oechel and G.R. Shaver, 2005: Arctic tundra and polar ecosystems. Arctic Climate Impact Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 243-351. [NPR, SRC]
Carmack, E.C. and R.W. Macdonald, 2002: Oceanography of the Canadian Shelf of the Beaufort Sea: a setting for marine life. Arctic, 55, 29-45. Clean
Chabot, M., 2004: Consumption and standards of living of the Québec Inuit: cultural permanence and discontinuities. Can. Rev. Sociol. Anthr., 41, 147-170. Clean
Chapin, F.S., III, G. Peterson, F. Berkes, T.V. Callaghan, P. Angelstam, M. Apps, C. Beier, C. Bergeron and Co-authors, 2004: Resilience and vulnerability of northern regions to social and environmental change. Ambio, 33, 342-347. [SRC]
Chapin, F.S., III, M. Berman, T.V. Callaghan, P. Convey, A.-S. Crepin, K. Danell, H. Ducklow, B. Forbes and Co-authors, 2005a: Polar systems. The Millennium Ecosystem Assessment, R. Hassan, R. Scholes and N. Ash, Eds., Island Press, Washington, District of Columbia, 717-743. [NPR, SRC]
Chapin, F.S., III, M. Sturm, M.C. Serreze, J.P. McFadden, J.R. Key, A.H. Lloyd, A.D. McGuire, T.S. Rupp and Co-authors, 2005b: Role of land-surface changes in Arctic summer warming. Science, 310, 657-660. [JoC]
Chapin, F.S., III, M. Hoel, S.R. Carpenter, J. Lubchenko, B. Walker, T.V. Callaghan, C. Folke, S.A. Levin, K.-G. Mäler, C. Nilsson, S. Barrett, F. Berkes, A.-S. Crépin, K. Danell and Co-authors, 2006: Building resilience to manage Arctic change. Science, 35, 198-202. [JoC, SRC]
Chartrand, J., K. Lysyshyn, R. Couture, S.D. Robinson and M.M. Burgess, 2002: Digital geotechnical borehole databases and viewers for Norman Wells and Tuktoyaktuk, Northwest Territories. Open File Report, 3912. Geological Survey of Canada, CD-Rom. [NPR]
Chernov, Y.I. and N.V. Matveyeva, 1997: Arctic ecosystems in Russia. Ecosystems of the World 3: Polar and Alpine Tundra, F.E. Wielgolaski, Ed., Elsevier, Amsterdam, 361-507. [NPR]
Christensen, T.R., T. Johansson, H.J. Akerman, M. Mastepanov, N. Malmer, T. Friborg, P. Crill and B.H. Svensson, 2004: Thawing sub-arctic permafrost: effects on vegetation and methane emissions. Geophys. Res. Lett., 31, L04501, doi: 10.1029/2003GL018680. [JoC]
Coleman, J.M. and O.K. Huh, 2004: Major World Deltas: A Perspective from Space. Coastal Studies Institute, Louisiana State University, Baton Rouge, Louisiana. http://www.geol.lsu.edu/WDD/PUBLICATIONS/introduction.htm. [NPR]
Community of Arctic Bay, 2005: Inuit Observations on Climate and Environmental Change: Perspectives from Arctic Bay, Nunavut. ITK, Nasivvik, NAHO, NTI, Ottawa, 57 pp. [NPR]
Constable, A.J., W.K. de la Mare, D.J. Agnew, I. Everson and D. Miller, 2000: Managing fisheries to conserve the Antarctic marine ecosystem: practical implementation of the Convention on the Conservation of Antarctic Marine Living Resources (CCAMLR). ICES J. Mar. Sci., 57, 778-791. Clean
Convey, P., P.J.A. Pugh, C. Jackson, A.W. Murray, C.T. Ruhland, F.S. Xiong and T.A. Day, 2002: Response of Antarctic terrestrial microarthropods to long-term climate manipulations. Ecology, 83, 3130-3140. Clean
Cook, A., A.J. Fox, D.G. Vaughan and J.G. Ferrigno, 2005: Retreating glacier-fronts on the Antarctic Peninsula over the last 50 years. Science, 22, 541-544. [JoC, SRC]
Cornelissen, J.H.C., T.V. Callaghan, J.M. Alatalo, A.E. Hartley, D.S. Hik, S.E. Hobbie, M.C. Press, C.H. Robinson and Co-authors, 2001: Global change and Arctic ecosystems: is lichen decline a function of increases in vascular plant biomass? J. Ecol., 89, 984-994. [SRC]
Corradi, C., O. Kolle, K. Walters, S.A. Zimov and E.-D. Schulze, 2005: Carbon dioxide and methane exchange of a north-east Siberian tussock tundra. Glob. Change Biol., 11, 1910-1925. [JoC]
Couture, R., S.D. Robinson and M.M. Burgess, 2000: Climate change, permafrost degradation, and infrastructure adaptation: preliminary results from a pilot community study in the Mackenzie Valley. Geological Survey of Canada, Current Research 2000-B2, 9 pp. Clean
Couture, R., S.D. Robinson and M.M. Burgess, 2001: Climate change, permafrost degradation and impacts on infrastructure: two case studies in the Mackenzie Valley. Proc., 54th Canadian Geotechnical Conference: An Earth Odyssey, Calgary, 908-915. [NPR]
Couture, R., S. Smith, S.D. Robinson, M.M. Burgess and S. Solomon, 2003: On the hazards to infrastructure in the Canadian north associated with thawing of permafrost. Geohazards 2003: 3rd Canadian Conference on Geotechnique and Natural Hazards, Edmonton. Canadian Geotechnical Society, Alliston, Ontario, 97-104. [NPR, PoC]
Crawford, R.M.M., 2004: Long-term plant survival at high latitudes. Bot. J. Scotland, 56, 1-23. Clean
Crawford, R.M.M., C.E. Jeffree and W.G. Rees, 2003: Paludification and forest retreat in northern oceanic environments. Ann. Bot. Lond., 91, 213-226. Clean
Curran, M.A.J., T.D. van Ommen, V.I. Morgan, K.L. Phillips and A.S. Palmer, 2003: Ice core evidence for Antarctic sea ice decline since the 1950s. Science, 302, 1203-1206. [JoC]
Curtis, T., S. Kvernmo and P. Bjerregaard, 2005: Changing living conditions, lifestyle and health. International Journal of Circumpolar Health, 64, 442-450. Clean
Dalen, L. and A. Hofgaard, 2005: Differential regional treeline dynamics in the Scandes Mountains. Arct. Antarct. Alp. Res., 37, 284-296. Clean
Davis, C.H., L. Yonghong, J.R. McConnell, M.M. Frey and E. Hanna, 2005: Snowfall-driven growth in Antarctic Ice Sheet mitigates recent sea-level rise. Science, 308, 1898-1901. [JoC]
De Angelis, H. and P. Skvarca, 2003: Glacier surge after ice shelf collapse. Science, 299, 1560-1562. [JoC]
de la Mare, W.K., 1997: Abrupt mid-twentieth-century decline in Antarctic sea-ice extent from whaling records. Nature, 389, 57-60. [JoC]
Derocher, A.E., N.J. Lunn and I. Stirling, 2004: Polar bears in a warming climate. Integr. Comp. Biol., 44, 163-176. [ARC]
Dietrich, R.A., Ed., 1981: Alaskan Wildlife Diseases. Institute of Arctic Biology, University of Alaska, Fairbanks. [NPR]
Domack, E., A. Leventer, S. Root, J. Ring, E. Williams, D. Carlson, E. Hirshorn, W. Wright, R. Gilbert and G. Burr, 2003: Marine sedimentary record of natural environmental variability. Antarctic Peninsula Climate Variability: Historical and Paleoenvironmental Perspectives, E. Domack, A. Leventer, A. Burnett, R. Bindschadler, P. Convey and M. Kirby, Eds., Antarctic Research Series 79, AGU, Washington, District of Columbia, 61-68. [NPR]
Domack, E., D. Duran, A. Leventer, S. Ishman, S. Doane, S. McCallum, D. Amblas, J. Ring, R. Gilbert and M. Prentice, 2005: Stability of the Larsen B ice shelf on the Antarctic Peninsula during the Holocene epoch. Nature, 436, 681-685. [JoC]
Doran, P.T., J.C. Priscu, W. Berry Lyons, J.E. Walsh, A.G. Fountain, D.M. McKnight, D.L. Moorhead, R.A. Virginia and Co-authors, 2002: Antarctic climate cooling and terrestrial ecosystem response. Nature, 415, 517-520. [JoC, SRC]
Dore, M. and I. Burton, 2001: The Costs of Adaptation to Climate Change in Canada: A Stratified Estimate by Sectors and Regions – Social Infrastructure. Environment Canada, Ottawa, 339 pp. [NPR, MoS, ARC]
Dowdeswell, J.A., 2006: Atmospheric science: the Greenland Ice Sheet and global sea-level rise. Science, 311, 963-964. [JoC]
Downey, D. and T. Fenge, Eds., 2003: Northern Lights Against POPs: Combating Toxic Threats in the Arctic. McGill–Queen’s University Press, Montreal, 347 pp. [NPR]
Drinkwater, K.F., 2005: The response of Atlantic cod (Gadus morhua) to future climate change. ICES J. Mar. Sci., 62, 1327-1337. [MoS]
Duhaime, G., 2004: Economic systems. The Arctic Human Development Report, N. Einarsson, J.N. Larsen, A. Nilsson and O.R. Young, Eds., Stefansson Arctic Institute, Akureyri, 69-84. [NPR]
Dye, D.G., 2002: Variability and trends in the annual snow-cover cycle in northern hemisphere land areas, 1972-2000. Hydrol. Process., 16, 3065-3077. Clean
Dyurgerov, M.B. and C.L. Carter, 2004: Observational evidence of increases in freshwater inflow to the Arctic Ocean. Arct. Antarct. Alp. Res., 36, 117-122. [ARC]
Eagles, P.F.J., 2004: Trends affecting tourism in protected areas. Proc., 2nd International Conference on Monitoring and Management of Visitor Flows in Recreational and Protected Areas, Rovaniemi, Finland: Policies, Methods and Tools for Visitor Management, T. Sievänen, J. Erkkonen, J. Jokimäki, J. Saarinen, S. Tuulentie and E. Virtanen, Eds., Working Papers of the Finnish Forest Research Institute 2, 18-26. [NPR]
Edwards, M. and A.J. Richardson, 2004: Impact of climate change on marine pelagic phenology and trophic mismatch. Nature, 430, 881-884. [JoC, ARC]
Einarsson, N., J.N. Larsen, A. Nilsson and O.R. Young, Eds., 2004: Arctic Human Development Report. Stefansson Arctic Institute, Akureyri, 242 pp. [NPR]
Ekstrom, G., M. Nettles and V.C. Tsai, 2006: Seasonality and increasing frequency of Greenland glacial earthquakes. Science, 311, 1756-1758. [JoC]
Elmhagen, B., M. Tannerfeldt, P. Verucci and A. Angerbjörn, 2000: The arctic fox (Alopex lagopus): an opportunistic specialist. J. Zool. Soc. Lond., 251, 139-149. Clean
Esper, J. and F.H. Schweingruber, 2004: Large-scale treeline changes recorded in Siberia. Geophys. Res. Lett., 31, L06202, doi:10.1029/2003GL019178. [JoC]
Eugster, W., W.R. Rouse, R.A. Pielke, J.P. McFadden, D.D. Baldocchi, T.G.F. Kittel, F.S. Chapin III and Co-authors, 2000: Land-atmosphere energy exchange in Arctic tundra and boreal forest: available data and feedbacks to climate. Glob. Change Biol., 6, S84-S115. [JoC]
Euskirchen, S.E., A.D. McGuire, D.W. Kicklighter, Q. Zhuang, J.S. Clein, R.J. Dargaville, D.G. Dye and Co-authors, 2006: Importance of recent shifts in soil thermal dynamics on growing season length, productivity, and carbon sequestration in terrestrial high-latitude ecosystems. Glob. Change Biol., 12, 731-750. [JoC]
FAUNMAP Working Group, 1996: Spatial response of mammals to late Quaternary environmental fluctuations. Science, 272, 1601-1606. [JoC]
Flanagan, K.M., E. McCauley and F.J. Wrona, 2006: Freshwater food webs control saturation through sedimentation. Glob. Change Biol., 12, 644-651. [JoC, SRC]
Foley, J.A., J.E. Kutzbach, M.T. Coe and S. Levis, 1994: Feedbacks between climate and boreal forests during the Holocene epoch. Nature, 371, 52-54. [JoC]
Forbes, D.L., 2005: Coastal erosion. Encyclopedia of the Arctic, M. Nutall, Ed., Routledge, New York and London, 391-393. [NPR, SRC]
Forchhammer, M. and D. Boertmann, 1993: The muskoxen Ovibos moschatus in north and northeast Greenland: population trends and the influence of abiotic parameters on population dynamics. Ecography, 16, 299-308. Clean
Fowbert, J.A. and R. Smith, 1994: Rapid population increases in native vascular plants in the Argentine Islands, Antarctic Peninsula. Arct. Alp. Res., 26, 290-296. Clean
Fox, A.J. and A.P.R. Cooper, 1998: Climate-change indicators from archival aerial photography of the Antarctic Peninsula. Ann. Glaciol., 27, 636-642. Clean
Fraser, W.R. and E.E. Hoffmann, 2003: A predator’s perspective on causal links between climate change, physical forcing and ecosystem response. Mar. Ecol- Prog. Ser., 265, 1-15. [MoS]
Freeman, M., 1996: Identity, health and social order. Human Ecology and Health: Adaptation to a Changing World, M.-L. Foller and L.O. Hansson, Eds., Gothenburg University, Gothenburg, 57-71. [NPR]
Freeman, M.M.R., 2000: Endangered Peoples of the Arctic. Greenwood Press, Connecticut, 278 pp. [NPR]
Frenot, Y., S.L. Chown, J. Whinam, P.M. Selkirk, P. Convey, M. Skotnicki and D.M. Bergstrom, 2005: Biological invasions in the Antarctic: extent, impacts and implications. Biol. Rev., 80, 45-72. Clean
Frey, K.E. and L.C. Smith, 2005: Amplified carbon release from vast West Siberian peatlands by 2100. Geophys. Res. Lett., 32, L09401, doi:10.1029/2004GL022025. [JoC]
Friborg, T., H. Soegaard, T.R. Christensen, C.R. Lloyd and N.S. Panikov, 2003: Siberian wetlands: where a sink is a source. Geophys. Res. Lett., 30, 2129, doi:10.1029/2003GL017797. [JoC]
Furgal, C., D. Martin and P. Gosselin, 2002: Climate change and health in Nunavik and Labrador: lessons from Inuit knowledge. The Earth is Faster Now: Indigenous Observations on Arctic Environmental Change, I. Krupnik and D. Jolly, Eds., ARCUS, Washington, District of Columbia, 266-300. [NPR, SRC]
Gauthier, G., J.-F. Giroux, A. Reed, A. Béchet and L. Bélanger, 2005: Interactions between land use, habitat use, and population increase in greater snow goose: what are the consequences for natural wetlands? Glob. Change Biol., 11, 856-868. [JoC]
Gedney, N., P.M. Cox, R.A. Betts, O. Boucher, C. Huntingford and P.A. Stott, 2006: Detection of a direct carbon dioxide effect in continental river runoff records. Nature, 439, 835-838. [JoC, ARC]
George, J.C., H.P. Huntington, K. Brewster, H. Eicken, D.W. Norton and R. Glenn, 2004: Observations on shorefast ice dynamics in arctic Alaska and the responses of the Inupiat hunting community. Arctic, 57, 363-374. Clean
Georgievsky, V.Y., I.A. Shiklomanov and A.L. Shalygin, 2005: Climate change impact on the water runoff on Lena river basin. Challenges of Ecological Meteorology and Climatology, G.V. Menzhulin, Ed., Nauka, St. Petersburg, 218-232. [NPR, ARC]
Gibson, J.J., T.D. Prowse and D.L. Peters, 2006: Partitioning impacts of climate and regulation on water level variability in Great Slave Lake. J. Hydrol., 329, 196-206. [SRC]
Gray, A., 1995: The indigenous movement in Asia. Indigenous Peoples of Asia, A. Gray, R.H. Barnes and B. Kingsbury, Eds., Association for Asian Studies, Ann Arbor, Michigan. [NPR]
Grebmeier, J.M., J.E. Overland, S.E. Moore, E.V. Farley, E.C. Carmack, L.W. Cooper, K.E. Frey, J.H. Helle and Co-authors, 2006: A major ecosystem shift in the northern Bering Sea. Science, 311, 1461-1464. [JoC]
Gregory, J.M., P. Huybrechts and S.C.B. Raper, 2004: Climatology: threatened loss of the Greenland ice-sheet. Nature, 428, 616. [JoC, ARC]
Hafrannsóknastofnunin, 2006: ?ættir úr vistfræ?i sjávar 2005 (Environmental Conditions in Icelandic Waters 2005. In Icelandic with English table headings and summaries). Hafrannsóknastofnunin (Marine Research Institute), 36 pp. [NPR]
Hamilton, L.C., B.C. Brown and R.O. Rasmussen, 2003: West Greenland’s cod-to-shrimp transition: local dimensions of climate change. Arctic, 56, 271-282. Clean
Handmer, J.W., S. Dovers and T.E. Downing, 1999: Societal vulnerability to climate change and variability. Mitigation and Adaptation Strategies for Climate Change, 4, 267-281. [ARC]
Harris, C., D. Vonder Mühll, K. Isaksen, W. Haeberli, J.L. Sollid, L. King, P. Holmlund, F. Dramis and Co-authors, 2003: Warming permafrost in European mountains. Global Planet. Change, 39, 215-225. [ARC]
Harris, L., 1990: Independent review of the state of the northern cod stock. Final Report of the Northern Cod Review Panel, Department of Supply and Services, Ottawa, 154 pp. [NPR]
Hassi, J., M. Rytkonen, J. Kotaniemi and H. Rintimaki, 2005: Impacts of cold climate on human heat balance, performance, illnesses and injuries in circumpolar areas. International Journal of Circumpolar Health, 64, 459-567. Clean
Hayley, D.W., 2004: Climate Change: An Adaptation Challenge for Northern Engineers. Association of Professional Engineers, Geologists and Geophysicists of Alberta, Alberta. http://www.apegga.org/members/publications/peggs/web01-04/expert.htm. [NPR]
Henttonen, H. and H. Wallgren, 2001: Small rodent dynamics and communities in the birch forest zone of northern Fennoscandia. Nordic Mountain Birch Forest Ecosystems, F.E. Wielgolaski, Ed., Man and the Biosphere Series. UNESCO, Paris, and Parthenon Publishing Group, New York and London, 261-278. [NPR]
Hild, C. and V. Stordhal, 2004: Human health and well-being. Arctic Human Development Report (AHDR), N. Einarsson, J.N. Larsen, A. Nilsson, O.R. Young, Eds., Steffanson Arctic Institute, Akureyri, 155-168. [NPR]
Hinkel, K.M., F.E. Nelson, A.F. Klene and J.H. Bell, 2003: The urban heat island in winter at Barrow, Alaska. Int. J. Climatol., 23, 1889-1905. [JoC, SRC]
Hinzman, L.D., N.D. Bettez, W.R. Bolton, F.S. Chapin, M.B. Dyurgerov, C.L. Fastie, B. Griffith, R.D. Hollister and Co-authors, 2005: Evidence and implications of recent climate change in northern Alaska and other Arctic regions. Climatic Change, 72, 251-298. [JoC, SRC]
Hobbie, J.E., B.J. Peterson, N. Bettez, L. Deegan, J. O’Brien, G.W. Kling and G.W. Kipphut, 1999: Impact of global change on biogeochemistry and ecosystems of arctic Alaska freshwaters. Polar Res., 18, 207-214. Clean
Høgda, K.A., S.R. Karlsen and H. Tømmervik, 2007: Changes in growing season in Fennoscandia 1982-1999. Arctic-Alpine Ecosystems and People in a Changing Environment, J.B. Orbaek, R. Kallenborn, I. Tombre, E.N. Hegseth, S. Falk-Petersen and A.H. Hoel, Eds., Springer-Verlag, Berlin, 71-84. [NPR, 2007]
Holland, M.M. and C.M. Bitz, 2003: Polar amplification of climate change in coupled models. Clim. Dynam., 21, 221-232. [JoC, MoS]
Hörnberg, S., 1995: Moose density related to occurrence and consumption of different forage species in Sweden. SLU Institution för Skogstaxering, Umeå. Rapporter–Skog, 58l, 34. Clean
Horsted, S.A., 2000: A review of the cod fisheries at Greenland, 1910-1995. Journal of Northwest Atlantic Fishery Science, 28, 1-112. Clean
Howat, I.M., I. Joughin, S. Tulaczyk and S. Gogineni, 2005: Rapid retreat and acceleration of Helheim glacier, east Greenland. Geophys. Res. Lett., 32, L22502, doi:10.1029/2005GL024737. [JoC]
Hughes-Hanks, J.M., L.G. Rickard, C. Panuska, J.R. Saucier, T.M. O’Hara, R.M. Rolland and L. Dehn, 2005: Prevalence of Cryptosporidium spp. and Giardia spp. in five marine mammal species. J. Parasitol., 91, 1357-1357. Clean
Huisink, M., J.J.W. De Moor, C. Kasse and T. Virtanen, 2002: Factors influencing periglacial fluvial morphology in the northern European Russian tundra and taiga. Earth Surf. Proc. Land., 27, 1223-1235. Clean
Huntington, H., 1998: Observations on the utility of the semi-directive interview for documenting traditional ecological knowledge. Arctic, 51, 237-242. Clean
Huntington, H., T.V. Callaghan, S. Fox and I. Krupnik, 2004: Matching traditional and scientific observations to detect environmental change: a discussion on arctic terrestrial ecosystems. Ambio, 33(S13), 18-23. [SRC]
Huntington, H., S. Foxand Co-authors, 2005: The changing Arctic: Indigenous perspectives. Arctic Climate Impact Assessment, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 61-98. [NPR]
Huntington, H.P., 1992: Wildlife Management and Subsistence Hunting in Alaska. Belhaven Press, London, 177 pp. [NPR]
Hylen, A., 2002: Fluctuations in abundance of northeast Arctic cod during the 20th century. 100 Years of Science under ICES: Papers from a Symposium held in Helsinki, 1-4 August 2000, E.D. Anderson, Ed., Marine Science Symposia 215, International Council for the Exploration of the Sea, 543-550. [NPR]
IAATO, 2005: Update on boot and clothing decontamination guidelines and the introduction and detection of diseases in Antarctic wildlife: IAATO’s perspective. Paper submitted by the International Association of Antarctica Tour Operators (IAATO) to the Antarctic Treaty Consultative Meeting (ATCM) XXVIII. IAATO, 10 pp. http://www.iaato.org/info.html. [NPR]
IAATO, 2006: IAATO Overview of Antarctic tourism 2005–2006. Information paper submitted by the International Association of Antarctica Tour Operators (IAATO) to the Antarctic Treaty Consultative Meeting (ATCM) XXIX. IAATO, 21 pp. http://www.iaato.org/info.html. [NPR]
ICARP II, 2006: Science Plan 8: terrestrial and freshwater biosphere and biodiversity. Proc., 2nd International Conference on Artic Research Planning (ICARP II), Copenhagen, 20 pp. [NPR]
ICES, 2005a: Report of the Arctic Fisheries Working Group, Murmansk, 19-28 April 2005. ICES CM 2005/ACFM:20. International Council for the Exploration of the Sea, Copenhagen, 564 pp. [NPR]
ICES, 2005b: Report of the Northwestern Working Group (NWWG) on 26 April-5 May 2005. ICES CM 2005/ACFM:21. International Council for the Exploration of the Sea, Copenhagen, 615 pp. [NPR]
ICES, 2006: Report of the Northwestern Working Group (NWWG). ICES CM 2006/ACFM:26. International Council for the Exploration of the Sea, Copenhagen, 612 pp. [NPR]
Ims, R.A. and E. Fuglei, 2005: Trophic interaction cycles in tundra ecosystems and the impact of climate change. BioScience, 554, 311-322. Clean
Instanes, A., O. Anisimov, L. Brigham, D. Goering, B. Ladanyi, J.O. Larsen and L.N. Khrustalev, 2005: Infrastructure: buildings, support systems, and industrial facilities. Arctic Climate Impact Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 907-944. [NPR, SRC]
Jefferies, R.L., R.H. Drent and J.P. Bakker, 2006: Connecting arctic and temperate wetlands and agricultural landscapes: the dynamics of goose populations in response to global change. Wetlands and Natural Resource Management, J.T. Verhoeven, B. Beltman, R. Bobbink and D.F. Whigham, Eds., Ecological Studies 190, Springer, Berlin, 293-312. [NPR, SRC]
Jensen, A.S., 1926: Indberetning av S/S Dana’s praktisk videndskabelige fiskeriundersøgelser ved Vestgrønland i 1925. Beretninger og Kundgørelser vedr. Styrelsen af Grønland, 2, 291-315. Clean
Jensen, A.S., 1939: Concerning a change of climate during recent decades in the Arctic and Subarctic regions, from Greenland in the west to Eurasia in the east, and contemporary biological and physical changes. Det. Kgl. Danske Videnskabernes Selskab. Biologiske Medd., 14, 1-77. Clean
Jernsletten, J.-L. and K. Klokov, 2002: Sustainable Reindeer Husbandry. University of Tromso, Tromso, 164 pp. http://www.reindeer-husbandry.uit.no/. [NPR]
Johannessen, O.M., K. Khvorostovsky, M.W. Miles and L.P. Bobylev, 2005: Recent ice-sheet growth in the interior of Greenland. Science, 310, 1013-1016. [JoC]
Johansson, T., N. Malmer, P.M. Crill, T. Friborg, J.H. Åkerman, M. Mastepanov and T.R. Christensen, 2006: Decadal vegetation changes in a northern peatland, greenhouse gas fluxes and net radiative forcing. Glob. Change Biol., 12, 2352-2369. [JoC, MoS]
Johnson, D., C.D. Campbell, D. Gwynn-Jones, J.A. Lee and T.V. Callaghan, 2002: Arctic soil microorganisms respond more to long-term ozone depletion than to atmospheric CO2. Nature, 416, 82-83. [JoC, SRC]
Jones, C.D. and D.C. Ramm, 2004: The commercial harvest of krill in the southwest Atlantic before and during the CCAMLR 2000 Survey. Deep-Sea Res. Pt. II, 51, 1421-1434. Clean
Juday, G.P., V. Barber, P. Duffy, H. Linderholm, S. Rupp, S. Sparrow, E. Vaganov and J. Yarie, 2005: Forests, land management and agriculture. Arctic Climate Impact Assessment ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 781-862. [NPR]
Kaplan, J.O., N.H. Bigelow, I.C. Prentice, S.P. Harrison, P.J. Bartlein, T.R. Christensen, W. Cramer, N.V. Matveyeva and Co-authors, 2003: Climate change and Arctic ecosystems. 2. Modeling, paleodata-model comparisons, and future projections. J. Geophys. Res., 108, 8171, doi:10.1029/2002JD002559. [JoC, MoS, ARC]
Kattsov, V.M., J.E. Walsh, W.L. Chapman, V.A. Govorkova, T.V. Pavlova and X. Zhang, 2007: Simulation and projection of Arctic freshwater budget components by the IPCC AR4 global climate models. J. Hydrometeorol., 8, 571-589. [JoC, MoS, SRC, 2007]
Khasnullin, V.I., A.M. Shurgaya, A.V. Khasnullina and E.V. Sevostoyanova, 2000: Cardiomyopathies in the North: Novosibirsk (in Russian). Siberian Division of Russian Academy of Medical Sciences, Siberia, 222 pp. [NPR]
Khrustalev, L.N., 2000: On the necessity of accounting for the effect of changing climate in permafrost engineering. Geocryological Hazards, L.S. Garagulia and E.D. Yershow, Eds., Kruk Publishers, Moscow, 238-247. [NPR]
King, J.C., 2003: Antarctic Peninsula climate variability and its causes as revealed by analysis of instrumental records. Antarctic Peninsula Climate Variability: Historical and Paleoenvironmental Perspectives. Antarctic Research Series 79, AGU, Washington, District of Columbia, 17-30. [NPR]
Klein, D., 1989: Subsistence hunting. Wildlife Production Systems: Economic Utilisation of Wild Ungulates, R.J. Hudson, K.R. Drew and L.M. Baskin, Cambridge University Press, Cambridge, 96-111. [NPR]
Klein, D., L.M. Baskin, L.S. Bogoslovskaya, K. Danell, A. Gunn, D.B. Irons, G.P. Kofinas, K.M. Kovacs and Co-authors, 2005: Management and conservation of wildlife in a changing Arctic. Arctic Climate Impact Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 597-648. [NPR]
Kofinas, G., G. Osherenko, D. Klein and B. Forbes, 2000: Research planning in the face of change: the human role in reindeer/caribou systems. Polar Res., 19, 3-22. Clean
Kokelj, S.V. and C.R. Burn, 2005: Near-surface ground ice in sediments of the Mackenzie Delta, Northwest Territories, Canada. Permafrost Periglac., 16, 291-303. Clean
Korhola, A., S. Sorvari, M. Rautio, P.G. Appleby, J.A. Dearing, Y. Hu, N. Rose, A. Lami and N.G. Cameron, 2002: A multi-proxy analysis of climate impacts on recent ontogeny of subarctic Lake Sannajärvi in Finnish Lapland. J. Paleolimnol., 1, 59-77. Clean
Koshida, G. and W. Avis, Eds., 1998: Canada Country Study: Climate Impacts and Adaptation. Vol. 7, National Sectoral Issues. Environment Canada, Toronto, 620 pp. [NPR]
Kovats, R.S., B. Menne, A.J. McMichael, C. Corvalan and R. Bertollini, 2000: Climate change and human health: impact and adaptation. WHO/SDE/OEH/00.4. World Health Organization, Geneva, 48 pp. [NPR, ARC]
Krabill, W., W. Abdalati, E. Frederick, S. Manizade, C. Martin, J. Sonntag, R. Swift, R. Thomas, W. Wright and J. Yungel, 2000: Greenland ice sheet: high-elevation balance peripheral thinning. Science, 289, 428-430. [JoC]
Krabill, W., E. Hanna, P. Huybrechts, W. Abdalati, J. Cappelen, B. Csatho, E. Frederick, S. Manizade and Co-authors, 2004: Greenland Ice Sheet: increased coastal thinning. Geophys. Res. Lett., 31, L24402, doi:10.1029/2004GL021533. [JoC]
Kraemer, L., J. Berner and C. Furgal, 2005: The potential impact of climate change on human exposure to contaminants in the Arctic. International Journal for Circumpolar Health, 64, 498-509. [SRC]
Krupnik, I., 1993: Arctic Adaptations: Native Whalers and Reindeer Herders of Northern Eurasia. University Press of New England, Lebanon, Pennsylvania, 355 pp. [NPR]
Krupnik, I. and D. Jolly, Eds., 2002: The Earth is Faster Now: Indigenous Observations of Arctic Environmental Change. Arctic Research Consortium of the United States, Fairbanks, Alaska, 356 pp. [NPR]
Kuhnlein, H.V., O. Receveur and H.M. Chan, 2001: Traditional food systems research with Canadian indigenous peoples. International Journal of Circumpolar Health, 60, 112-122. Clean
Kutz, S.J., E.P. Hoberg, J. Nishi and L. Polley, 2002: Development of the musk ox lungworm Umingmakstrongylus pallikuukensis (Protostrongylidae) in gastropods in the Arctic. Can. J. Zool., 80, 1977-1985. Clean
Lafortune, V., C. Furgal, J. Drouin, T. Annanack, N. Einish, B. Etidloie, M. Qiisiq, P. Tookalook and Co-authors, 2004: Climate change in northern Québec: access to land and resource issues. Project report. Kativik Regional Government, Kuujjuaq, Québec. [NPR, SRC]
Laherre, J., 2001: Estimates of oil reserves. Preprints EMF/IEA/IEW Meeting, International Institute for Applied Systems Analysis (IIASA), Laxenburg, 92 pp. [NPR]
Lantuit, H. and W. Pollard, 2003: Remotely sensed evidence of enhanced erosion during the twentieth century on Herschel Island, Yukon Territory. Berichte zur Polar- und Meeresforschung, 443, 54-59. Clean
Lemke, P., J. Ren, R. Alley, I. Allison, J. Carrasco, G. Flato, Y. Fujii, G. Kaser, P. Mote, R. Thomas and T. Zhang, 2007: Observations: change in snow, ice and frozen ground. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor and H.L. Miller, Eds., Cambridge University Press, Cambridge, 337-384. [NPR, ARC, 2007]
Lesack, L.F.W., P. Marsh and R.E. Hecky, 1998: Spatial and temporal dynamics of major solute chemistry along Mackenzie Delta lakes. Limnol. Oceanogr., 43, 1530-1543. Clean
Lewis, E.L., E.P. Jones, P. Lemke, T.D. Prowse and P. Wadhams, 2000: The Freshwater Budget of the Arctic Ocean. Kluwer Academic, Dordrecht, 623 pp. [NPR, SRC]
Lilly, G.R., P.A. Shelton, J. Brattey, N. Cadigan, B.P. Healey, E.F. Murphy, D. Stanbury and N. Chen, 2003: An assessment of the cod stock in NAFO Divisions 2J+3KL in February 2003. 2003/023 Department of Fisheries and Oceans Stock Assessment Secretariat, 157 pp. [NPR]
Lindgren, E. and R. Gustafson, 2001: Tick-borne encephalitis in Sweden and climate change. Lancet, 358, 16-18. Clean
Lloyd, A.H., T.S. Rupp, C.L. Fastie and A.M. Starfield, 2003: Patterns and dynamics of treeline advance on the Seward Peninsula, Alaska. J. Geophys. Res., 108, 8161, doi:10.1029/2001JD000852. [JoC]
Loeng, H., K. Brander, E. Carmack, S. Denisenko, K. Drinkwater, B. Hansen, K. Kovacs, P. Livingston, F. McLaughlin and E. Sakshaug, 2005: Marine systems. Arctic Climate Impact Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 453-538. [NPR, ARC]
Lonergan, S., R. Difrancesco and M.-K. Woo, 1993: Climate change and transportation in northern Canada: an integrated impact assessment. Climatic Change, 24, 331-351. [JoC]
Luckman, A., T. Murray, R. de Lange and E. Hanna, 2006: Rapid and synchronous ice-dynamic changes in East Greenland. Geophys. Res. Lett., 33, L03503, doi:10.1029/2005GL025428. [JoC]
MacDonald, G.M., A.A. Velichko, C.V. Kremenetski, C.K. Borisova, A.A. Goleva, A.A. Andreev, L.C. Cwynar, R.T. Riding and Co-authors, 2000: Holocene tree line history and climate change across northern Eurasia. Quaternary Res., 53, 302-311. [ARC]
MacDonald, R., T. Harner, J. Fyfe, H. Loeng and T. Weingartner, 2003: Influence of global change on contaminant pathways to, within and from the Arctic. Arctic Monitoring and Assessment Programme, AMAP Assessment 2002, Oslo, 65 pp. [NPR]
Malmer, N., T. Johansson, M. Olsrud and T.R. Christensen, 2005: Vegetation, climatic changes and net carbon sequestration. Glob. Change Biol., 11, 1895-1909. [JoC]
Marsh, P. and L.F.W. Lesack, 1996: The hydrologic regime of perched lakes in the Mackenzie Delta: potential responses to climate change. Limnol. Oceanogr., 41, 849-856. Clean
Marsh, P., L.F.W. Lesack and A. Roberts, 1999: Lake sedimentation in the Mackenzie Delta, NWT. Hydrol. Process., 13, 2519-2536. Clean
Marshall, G.J., P.A. Stott, J. Turner, W.M. Connolley, J.C. King and T.A. Lachlan-Cope, 2004: Causes of exceptional atmospheric circulation changes in the Southern Hemisphere. Geophys. Res. Lett., 31, L14205, doi:10.1029/2004 GL019952. [JoC]
Marshall, G.J., A. Orr, N.P.M. van Lipzig and J.C. King, 2006: The impact of a changing southern hemisphere annular mode on Antarctic peninsula summer temperatures. J. Climate, 19, 5388-5404. [JoC]
Martin, D., D. Belanger, P. Gosselin, J. Brazeau, C. Furgal and S. Dery, 2005: Climate change, drinking water, and human health in Nunavik: adaptation strategies. Final Report submitted to the Canadian Climate Change Action Fund, Natural Resources Canada, Quebec, 111 pp. [NPR, SRC]
Maslin, M., 2004: Gas Hydrates: A Hazard for the 21st Century. Issues in Risk Science 3, Benfield Hazard Research Centre, London, 22 pp. [NPR]
Matveyeva, N.V. and Y. Chernov, 2000: Zonation in Plant Cover of the Arctic. Russian Academy of Sciences, Moscow, 219 pp. [NPR]
McBean, G., G. Alekseev, D. Chen, E. Førland, J. Fyfe, P.Y. Groisman, R. King, H. Melling, R. Vose and P.H. Whitfield, 2005: Arctic climate: past and present. Arctic Climate Impacts Assessment (ACIA), C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 21-60. [NPR, ARC]
McCarthy, J.J., M. Long Martello, R. Corell, N. Eckley Selin, S. Fox, G. Hovelsrud-Broda, S.D. Mathiesen, C. Polsky, H. Selin and N.J.C. Tyler, 2005: Climate change in the context of multiple stressors and resilience. Arctic Climate Impact Assessment (ACIA), C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 945-988. [NPR]
McClelland, J.W., R.M. Holmes and B.J. Peterson, 2004: Increasing river discharge in the Eurasian Arctic: consideration of dams, permafrost thaw, and fires as potential agents of change. J. Geophys. Res., 109, D18102, doi:10.1029/2004JD004583. [JoC]
McDonald, K.C., J.S. Kimball, E. Njoku, R. Zimmermann and M. Zhao, 2004: Variability in springtime thaw in the terrestrial high latitudes: monitoring a major control on the biospheric assimilation of atmospheric CO2 with spaceborne microwave remote sensing. Earth Interactions, 8, 1-23. Clean
McGuire, A.D., J.S. Clein, J.M. Melillo, D.W. Kicklighter, R.A. Meier, C.J. Vorosmarty and M.C. Serreze, 2000: Modeling carbon responses of tundra ecosystems to historical and projected climate: sensitivity of pan-Arctic carbon storage to temporal and spatial variation in climate. Glob. Change Biol., 6, S141-S159. [JoC, MoS]
McGuire, A.D., F.S. Chapin III, C. Wirth, M. Apps, J. Bhatti, T.V. Callaghan, T.R. Christensen, J.S. Clein, M. Fukuda, T. Maximov, A. Onuchin, A. Shvidenko and E. Vaganov, 2007: Responses of high latitude ecosystems to global change: potential consequences for the climate system. Terrestrial Ecosystems in a Changing World, J.G. Canadell, D.E. Pataki and L.F. Pitelka, Eds., Springer, London, 297-310. [NPR, SRC, 2007]
Meehl, G.H., T.F. Stocker, W.D. Collins, P. Friedlingstein, A.T. Gaye, J.M. Gregory, A. Kito, R. Knutti, J.M. Murphy, A. Noda, S.C.B. Raper, I.G. Watterson, A.J. Weaver and Z.-C. Zhao, 2007: Global climate projections. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor and H.L. Miller, Eds., Cambridge University Press, Cambridge, 747-846. [NPR, ARC, 2007]
Mehlum, F., 1999: Adaptation in arctic organisms to a short summer season. The Ecology of the Tundra in Svalbard, S.A. Bengtsson, F. Mehlum and T. Severinsen, Eds., Norsk Polarinstitutt Meddeleser, 161-169. [NPR]
Meredith, M.P. and J.C. King, 2005: Rapid climate change in the ocean west of the Antarctic peninsula during the second half of the 20th century. Geophys. Res. Lett., 32, L19604, doi:10.1029/2005GL024042. [JoC]
Messner, T., 2005: Environmental variables and the risk of disease. International Journal for Circumpolar Health, 64, 523-533. Clean
Miller, J.R. and G.L. Russell, 2000: Projected impact of climate change on the freshwater and salt budgets of the Arctic Ocean by a global climate model. Geophys. Res. Lett., 27, 1183-1186. [JoC, MoS]
Mokhov, I.I., V.A. Semenov and V.C. Khone, 2003: Estimates of possible regional hydrologic regime changes in the 21st century based on global climate models. Izvestiya Rossijskoi Academii Nauk (Proceedings of the Russian Academy of Sciences), 39, 130-144. [MoS]
Morrison, R.I.G., Y. Aubry, R.W. Butler, G.W. Beyresbergen, G.M. Donaldson, C.L. Gratto-Trevor, P.W. Hicklin, V.H. Johnston and R.K. Ross, 2001: Declines in North American shorebird populations. Wader Study Group Bulletin, 94, 34-38. Clean
Morrison, R.I.G., K.R. Ross and L.J. Niles, 2004: Declines in wintering populations of red knots in southern South America. Condor, 106, 60-70. Clean
Murphy, E.J., A. Clarke, C. Symon and J. Priddle, 1995: Temporal variation in Antarctic sea-ice: analysis of a long term fast ice record from the South Orkney Islands. Deep-Sea. Res., 42, 1045-1062. Clean
Nakićenović , N., J. Alcamo, G. Davis, B. de Vries, J. Fenhann, S. Gaffin, K. Gregory, A. Grübler, T.Y. Jung, T. Kram, E. Lebre la Rovere, L. Michaelis, S. Mori, T. Morita, W. Pepper, H. Pitcher, L. Price, K. Riahi, A. Roehrl, H.H. Rogner, A. Sankovski, M. Schlesinger, P. Shukla, S. Smith, R. Swart, S. van Rooijen, N. Victor and Z. Dadi, Eds., 2000: Emissions Scenarios: A Special Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, 599 pp. [NPR, ARC]
Nayha, S., 2005: Environmental temperature and mortality. International Journal of Circumpolar Health, 64, 451-458. Clean
Nellemann, C., L. Kullerud, I. Vistnes, B.C. Forbes, E. Husby, G.P. Kofinas, B.P. Kaltenborn, J. Rouaud and Co-authors, 2001: GLOBIO: global methodology for mapping human impacts on the biosphere. UNEP/DEWA/TR.01-3, United Nations Environment Programme, 47 pp. [NPR]
Nelson, F.E., 2003: (Un)frozen in time. Science, 299, 1673-1675. [JoC, SRC]
Nelson, F.E., O.A. Anisimov and N.I. Shiklomanov, 2001: Subsidence risk from thawing permafrost. Nature, 410, 889-890. [JoC, SRC]
Nemani, R.R., C.D. Keeling, H. Hashimoto, W.M. Jolly, S.C. Piper, C.J. Tucker, R.B. Myneni and S.W. Running, 2003: Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science, 300, 1560-1563. [JoC, ARC]
Nickels, S., C. Furgal, J. Castelden, P. Moss-Davies, M. Buell, B. Armstrong, D. Dillon and R. Fonger, 2002: Putting the human face on climate change through community workshops. The Earth is Faster Now: Indigenous Observations of Arctic Environmental Change, I. Krupnik and D. Jolly, Eds., ARCUS, Washington, District of Columbia, 300-344. [NPR, SRC]
Nikiforov, S.L., N.N. Dunaev, S.A. Ogorodov and A.B. Artemyev, 2003: Physical geographic characteristics. The Pechora Sea: Integrated Research. MOPE, Moscow, 502 pp. (In Russian). [NPR]
Nuttall, M., 2000a: The Arctic is Changing. Stefansson Arctic Institute, Akureyri, 11 pp. http://www.thearctic.is/PDF/The%20Arctic%20is%20changing.pdf. [NPR, SRC]
Nuttall, M., 2000b: Indigenous peoples, self-determination, and the Arctic environment. The Arctic: Environment, People, Policy, M. Nuttall and T.V. Callaghan, Eds., Harwood Academic, The Netherlands, 377-409. [NPR, SRC]
Nuttall, M., F. Berkes, B. Forbes, G. Kofinas, T. Vlassova and G. Wenzel, 2005: Hunting, herding, fishing and gathering: indigenous peoples and renewable resource use in the Arctic. Arctic Climate Impacts Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 649-690. [NPR, SRC]
Oechel, W.C., G.L. Vourlitis, S.J. Hastings, R.C. Zulueta, L. Hinzman and D. Kane, 2000: Acclimation of ecosystem CO2 exchange in the Alaskan Arctic in response to decadal climate warming. Nature, 406, 978-981. [JoC]
Ogorodov, S.A., 2003: Coastal dynamics in the Pechora Sea under technogenic impact. Berichte zur Polar- und Meeresforschung, 443, 74-80. Clean
Orr, J.C., V.J. Fabry, O. Aumont, L. Bopp, S.C. Doney, R.A. Feely, A. Gnanadesikan, N. Gruber and Co-authors, 2005: Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature, 437, 681-686. [PoC, JoC]
Paine, R., 1994: Herds of the Tundra: A Portrait of Saami Reindeer Pastoralism. Smithsonian Institution Press, Washington, District of Columbia, 242 pp. [NPR]
Parkinson, A. and J. Butler, 2005: Impact of climate change on infectious diseases in the Arctic. International Journal for Circumpolar Health, 64, 478. Clean
Parkinson, C.L., 2002: Trends in the length of the Southern Ocean sea ice season, 1979-1999. Ann. Glaciol., 34, 435-440. Clean
Parkinson, C.L., 2004: Southern Ocean sea ice and its wider linkages: insights revealed from models and observations. Antarct. Sci., 16, 387-400. [MoS]
Pavlov, A.V. and N.G. Moskalenko, 2002: The thermal regime of soils in the north of western Siberia. Permafrost Periglac., 13, 43-51. Clean
Pavlov, P., J.I. Svendsen and S. Indrelid, 2001: Human presence in the European Arctic nearly 40,000 years ago. Nature, 413, 64-67. [JoC]
Payette, S., M. Eronen and J.J.P. Jasinski, 2002: The circumpolar tundra-taiga interface: late Pleistocene and Holocene changes. Ambio, 12, 15-22. Clean
Payne, A.J., A. Vieli, A. Shepherd, D.J. Wingham and E. Rignot, 2004: Recent dramatic thinning of largest West Antarctic ice stream triggered by oceans. Geophys. Res. Lett., 31, L23401, doi:10.1029/2004GL021284. [JoC]
Peck, L.S., 2005: Prospects for surviving climate change in Antarctic aquatic species. Frontiers in Zoology, 2, doi:10.1186/1742-9994-2-9. [NPR]
Peck, L.S., P. Convey and D.K.A. Barnes, 2006: Environmental constraints on life histories in Antarctic ecosystems: tempos, timings and predictability. Biol. Rev., 81, 75-109. [MoS]
Peters, D.L., T.D. Prowse, A. Pietroniro and R. Leconte, 2006: Establishing the flood hydrology of the Peace–Athabasca Delta, northern Canada. Hydrol. Process., 20, 4073-4096. [SRC]
Peterson, B.J., R.M. Holmes, J.W. McClelland, C.J. Vorosmarty, R.B. Lammers, A.I. Shiklomanov, I.A. Shiklomanov and S. Rahmstorf, 2002: Increasing river discharge to the Arctic Ocean. Science, 298, 2172-2173. [PoC, JoC, ARC]
Peterson, B.J., J. McClelland, R. Curry, R.M. Holmes, J.E. Walsh and K. Aagaard, 2006: Trajectory shifts in the arctic and subarctic freshwater cycle. Science, 313, 1061-1066. [JoC, SRC]
Pienitz, R., M.S.V. Douglas and J.P. Smol, 2004: Long-Term Environmental Change in Arctic and Antarctic Lakes. Vol. 8. Springer Verlag, 562 pp. [NPR]
Polyakov, I.V., G.V. Alekseev, R.V. Bekryaev, U. Bhatt, R.L. Colony, M.A. Johnson, V.P. Karklin, A.P. Makshtas, D. Walsh and A.V. Yulin, 2002: Observationally based assessment of polar amplification of global warming. Geophys. Res. Lett., 29, 1878. doi:10.1029/2001GL011111. [JoC]
Prowse, T.D. and S. Beltaos, 2002: Climatic control of river-ice hydrology: a review. Hydrol. Process., 16, 805-822. [SRC]
Prowse, T.D., D.L. Peters, S. Beltaos, A. Pietroniro, L. Romolo, J. Töyrä and R. Leconte, 2002: Restoring ice-jam floodwater to a drying delta ecosystem. Water Int., 27, 58-69. [SRC]
Prowse, T.D., F.J. Wrona and G. Power, 2004: Dams, reservoirs and flow regulation. Threats to Water Availability in Canada. National Water Resource Institute, Scientific Assessment Report No. 3, Environment Canada, Ottawa, 9-18. [NPR, SRC]
Prowse, T.D., F.J. Wrona, J. Reist, J.J. Gibson, J.E. Hobbie, L.M.J. Levesque and W.F. Vincent, 2006a: Climate change effects on hydroecology of arctic freshwater ecosystems. Ambio, 35, 347-358. [SRC]
Prowse, T.D., F.J. Wrona, J.D. Reist, J.J. Gibson, J.E. Hobbie, L. Levesque and W.F. Vincent, 2006b: Historical changes in arctic freshwater ecosystems. Ambio, 35, 339-346. [SRC]
Pudsey, C.J. and J. Evans, 2001: First survey of Antarctic sub-ice shelf sediments reveals mid-Holocene ice shelf retreat. Geology, 29, 787-790. Clean
Quayle, W.C., L.S. Peck, H. Peat, J.C. Ellis-Evans and P.R. Harrigan, 2002: Extreme responses to climate change in Antarctic lakes. Science, 295, 645-645. [JoC]
Quayle, W.C., P. Convey, L.S. Peck, J.C. Ellis-Evans, H.G. Butler and H.J. Peat, 2003: Ecological responses of maritime Antarctic lakes to regional climate change. Antarctic Peninsula Climate Variability: Historical and Palaeoenvironmental Perspectives, E. Domack, A. Leventer, A. Burnett, R. Bindschadler, P. Convey and M. Kirby, Eds., American Geophysical Union, Washington, District of Columbia, 159-170. [NPR]
Rachold, V., M.N. Grigoriev, F.E. Aré, S. Solomon, E. Reimnitz, H. Kassens and M. Antonov, 2000: Coastal erosion vs. riverine sediment discharge in the Arctic shelf seas. Geologisches Rundschau (Int. J. Earth Sci.), 89, 450-460. [PoC, JoC]
Rasumov, S.O., 2001: Thermoerosion modelling of ice-rich Arctic coast in stationary climatic conditions (in Russian). Kriosfera Zemli, 5, 50-58. [MoS]
Reist, J., F.J. Wrona, T.D. Prowse, M. Power, J.B. Dempson, R. Beamish, J.R. King, T.J. Carmichael and C.D. Sawatzky, 2006a: General effects of climate change effects on arctic fishes and fish populations. Ambio, 35, 370-380. [SRC]
Reist, J.D., F.J. Wrona, T.D. Prowse, J.B. Dempson, M. Power, G. Koeck, T.J. Carmichael, C.D. Sawatzky, H. Lehtonen and R.F. Tallman, 2006b: Effects of climate change and UV radiation on fisheries for arctic freshwater and anadromous species. Ambio, 35, 402-410. [SRC]
Reist, J.D., F.J. Wrona, T.D. Prowse, M. Power, J.B. Dempson, J.R. King and R.J. Beamish, 2006c: An overview of effects of climate change on arctic freshwater and anadromous fisheries. Ambio, 35, 381-387. [SRC]
Richardson, A.J. and D.S. Schoeman, 2004: Climate impact on ecosystems in the northeast Atlantic. Science, 305, 1609-1612. [JoC]
Riebesell, U., I. Zondervan, B. Rost, P.D. Tortell, R.E. Zeebe and F.M. Morel, 2000: Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature, 407, 364-367. [JoC]
Riedlinger, D. and F. Berkes, 2001: Contributions of traditional knowledge to understanding climate change in the Canadian Arctic. Polar Rec., 37, 315-328. Clean
Rignot, E. and P. Kanagaratnam, 2006: Changes in the velocity structure of the Greenland ice sheet. Science, 311, 986-990. [JoC]
Rignot, E., G. Casassa, S. Gogineni, P. Kanagaratnam, W. Krabill, H. Pritchard, A. Rivera, R. Thomas and D. Vaughan, 2005: Recent ice loss from the Fleming and other glaciers,Wordie Bay, West Antarctic Peninsula. Geophys. Res. Lett., 32, L07502, doi:10.1029/2004GL021947. [JoC, SRC]
Rinnan, R., M.M. Keinänen, A. Kasurinen, J. Asikainen, T.K. Kekki, T. Holopainen, H. Ro-Poulsen, T.N. Mikkelsen and Co-authors, 2005: Ambient ultraviolet radiation in the Arctic reduces root biomass and alters microbial community composition but has no effects on microbial biomass. Glob. Change Biol., 11, 564-574. [JoC]
Robinson, S.A., J. Wasley and A.K. Tobin, 2003: Living on the edge: plants and global change in continental and maritime Antarctica. Glob. Change Biol., 9, 1681-1717. [JoC]
Romanovsky, V.E., M. Burgess, S. Smith, K.Yoshikawa and J. Brown, 2002: Permafrost temperature records: indicators of climate change. EOS Transactions, 83, 589-594. Clean
Rose, G.A., 2004: Reconciling overfishing and climate change with stock dynamics of Atlantic cod (Gadus morhua) over 500 years. Can. J. Fish. Aquat. Sci., 61, 1553-1557. [SRC]
Rose, G.A. and R.L. O’Driscoll, 2002: Capelin are good for cod: can Newfoundland cod stocks recover without capelin? ICES J. Mar. Sci., 5938, 1026. [SRC]
Rose, G.A., B. de Young, D.W. Kulka, S.V. Goddard and G.L. Fletcher, 2000: Distribution shifts and overfishing the northern cod (Gadus morhua): a view from the ocean. Can. J. Fish. Aquat. Sci., 57, 644-664. [SRC]
Roura, R., 2004: Monitoring and remediation of hydrocarbon contamination at the former site of Greenpeace’s World Park Base, Cape Evans, Ross Island, Antarctica. Polar Res., 40, 51-67. Clean
Royal Society, 2005: Ocean acidification due to increasing atmospheric carbon dioxide. Policy Document 12/05, The Royal Society, London, 60 pp. [NPR]
Ruhland, K.M. and J.P. Smol, 2002: Freshwater diatoms from the Canadian arctic treeline and development of paleolimnological inference models. J. Phycol., 38, 429-264. [MoS]
Ruhland, K.M., A. Priesnitz and J.P. Smol, 2003: Paleolimnological evidence from diatoms for recent environmental changes in 50 lakes across Canadian Arctic treeline. Arct. Antarct. Alp. Res., 35, 110-123. Clean
Russell, D.E., G. Kofinas and B. Griffith, 2002: Barren-ground caribou calving ground workshop. Canadian Wildlife Services, Ottawa, 47 pp. [NPR]
Sakshaug, E., 2003: Primary and secondary production in Arctic seas. The Organic Carbon Cycle in the Arctic Ocean, R. Stein and R.W. Macdonald, Eds., Springer, Berlin, 57-81. [NPR]
Sakshaug, E., A. Bjorge, B. Gulliksen, H. Loeng and F. Mehlum, 1994: Structure, biomass distribution and energetics of the pelagic ecosystem in the Barents Sea: a synopsis. Polar Biol., 14, 405-411. Clean
Sazonova, T.S., V.E. Romanovsky, J.E. Walsh and D.O. Sergueev, 2004: Permafrost dynamics in the 20th and 21st centuries along the East Siberian transect. J. Geophys. Res.–Atmos., 109, D01108, doi:10.1029/2003JD003680. [SRC]
Scambos, T.A., J.A. Bohlander, C.A. Shuman and P. Skvarca, 2004: Glacier acceleration and thinning after ice shelf collapse in the Larsen B embayment, Antarctica. Geophys. Res. Lett., 31, L18402, doi:10.1029/2004GL020670. [JoC]
Schopka, S.A., 1994: Fluctuations in the cod stock off Iceland during the twentieth century in relation to changes in the fisheries and environment. ICES Mar. Sci. Symp., 198, 175-193. Clean
Seebacher, F., W. Davison, C.J. Lowe and C.E. Franklin, 2005: The falsification of the thermal specialization paradigm: compensation for elevated temperatures in Antarctic fishes. Biol. Lett., 1, 151-154. Clean
Serreze, M.C. and J.A. Francis, 2006: The Arctic amplification debate. Climatic Change, 76, 241-264. [JoC]
Serreze, M.C., D.H. Bromwich, M.P. Clark, A.J. Etringer, T. Zhang and R. Lammers, 2003: Large-scale hydro-climatology of the terrestrial Arctic drainage system. J. Geophys. Res., 108, 8160, doi:10.1029/2001JD000919. [JoC]
Shelton, P.A., A.F. Sinclair, G.A. Chouinard, R. Mohn and D.E. Duplisea, 2006: Fishing under low productivity conditions is further delaying recovery of Northwest Atlantic cod (Gadus morhua).Can. J. Fish. Aquat. Sci., 63, 235-238. Clean
Shepherd, A., D.J. Wingham and E. Rignot, 2004: Warm ocean is eroding West Antarctic Ice Sheet. Geophys. Res. Lett., 31, L23402, doi:10.1029\2004G L021106. [JoC]
Shiklomanov, I.A., A.I. Shiklomanov, R.B. Lammers, B.J. Peterson and C.J. Vorosmarty, 2000: The dynamics of river water inflow to the Arctic Ocean. The Freshwater Budget of the Arctic Ocean, E.L. Lewis, E.P. Jones, T.D. Prowse and P. Wadhams, Eds., Kluwer Academic, Dordrecht, 281-296. [NPR, ARC]
Sirotenko, O.D., H.V. Abashina and V.N. Pavlova, 1997: Sensitivity of the Russian agriculture to changes in climate, CO2 and tropospheric ozone concentrations and soil fertility. Climatic Change, 36, 217-232. [JoC]
Sitch, S., B. Smith, I.C. Prentice, A. Arneth, A. Bondeau, W. Cramer, J.O. Kaplan, S. Levis, W. Lucht, M.T. Sykes, K. Thonicke and S. Venevsky, 2003: Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Change Biol., 9, 161-185. [JoC, MoS, ARC]
Sitch, S., A.D. McGuire, J. Kimball, N. Gedney, J. Gamon, R. Engstrom, A. Wolf, Q. Zhuang, J. Clein and K.C. McDonald, 2007: Assessing the carbon balance of circumpolar arctic tundra using remote sensing and process modeling. Ecol. Appl., 17, 213-234. [MoS, 2007]
Slayback, D.A., J.E. Pinzon, S.O. Los and C.J. Tucker, 2003: Northern hemisphere photosynthetic trends 1982–99. Glob. Change Biol., 9, 1-15. [JoC]
Sloan, E.D., Jr, 2003: Fundamental principles and applications of natural gas hydrates. Nature, 426, 353-359. [JoC]
Smith, L.C., 2000: Time-trends in Russian Arctic river ice formation and breakup: 1917–1994. Phys. Geogr., 21, 46-56. Clean
Smith, L.C., Y. Sheng, G.M. MacDonald and L.D. Hinzman, 2005: Disappearing Arctic lakes. Science, 308, 1429. [JoC]
Smith, N.V., S.S. Saatchi and J.T. Randerson, 2004: Trends in high northern latitude soil freeze and thaw cycles from 1988 to 2002. J. Geophys. Res., 109, D12101, doi:10.1029/2003JD004472. [JoC]
Smith, O.P., 2002: Coastal erosion in Alaska. Berichte zur Polar- und Meeresforschung, 413, 65-68. Clean
Smith, R.C., W.R. Fraser, S.E. Stammerjohn and M. Vernet, 2003: Palmer long-term ecological research on the Antarctic marine ecosystem. Antarctic Peninsula Climate Variability: Historical and Paleoenvironmental Perspectives, E. Domack, A. Leventer, A. Burnett, R. Bindschadler, P. Convey and M. Kirby, Eds., Antarctic Research Series 79, AGU, Washington, District of Columbia, 131-144. [NPR]
Smith, S.L. and M.M. Burgess, 2004: Sensitivity of Permafrost to Climate Warming in Canada. Geological Survey of Canada Bulletin No. 579, 24 pp. [NPR]
Smith, S.L., M.M. Burgess and J.A. Heginbottom, 2001: Permafrost in Canada, a challenge to northern development. A Synthesis of Geological Hazards in Canada, G.R. Brooks, Ed., Geological Survey of Canada Bulletin 548, 241-264. [NPR]
Smol, J.P., A.P. Wolfe, H.J.B. Birks, M.S.V. Douglas, V.J. Jones, A. Korhola, R. Pienitz, K. Ruhland and Co-authors, 2005: Climate-driven regime shifts in the biological communities of arctic lakes. P. Natl Acad. Sci. USA, 102, 4397-4402. [PoC, JoC]
Snape, I., M.J. Riddle, D.M. Filler and P.J. Williams, 2003: Contaminants in freezing ground and associated ecosystems: key issues at the beginning of the new millennium. Polar Rec., 39, 291-300. Clean
Solomon, S.M., 2005: Spatial and temporal variability of shoreline change in the Beaufort-Mackenzie region, Northwest Territories, Canada. Geo-Marine Lett., 25, 127-137. [PoC, JoC]
Stieglitz, M., S.J. Dery, V.E. Romanovsky and T.E. Osterkamp, 2003: The role of snow cover in the warming of arctic permafrost. Geophys. Res. Lett., 30, 1721, doi:10.1029/2003GL017337. [JoC]
Stirling, I. and C.L. Parkinson, 2006: Possible effects of climate warming on selected populations of polar bears (Ursus maritimus) in the Canadian Arctic. Arctic, 59, 262-275. Clean
Stirling, I., N.J. Lunn and J. Iacozza, 1999: Long-term trends in the population ecology of polar bears in western Hudson Bay in relation to climate change. Arctic, 52, 294-306. [ARC]
Stone, J.O., G.A. Balco, D.E. Sugden, M.W. Caffee, L.C. Sass III, S.G. Cowdery and C. Siddoway, 2003: Holocene deglaciation of Marie Byrd Land, West Antarctica. Science, 299, 99-102. [JoC]
Strathdee, A.T., J.S. Bale, W.C. Block, S.J. Coulson, I.D. Hodkinson and N. Webb, 1993: Effects of temperature on a field population of Acyrthosiphon svalbardicum (Hemiptera: Aphidae) on Spitsbergen. Oecologia, 96, 457-465. Clean
Stroud, D.A., N.C. Davidson, R. West, D.A. Scott, L. Haanstra, O. Thorup, B. Ganter and S. Delany, 2004: Status of migratory wader population in Africa and Western Eurasia in the 1990s. International Wader Studies, 15, 1-259. Clean
Sturm, M., C. Racine and K. Tape, 2001: Increasing shrub abundance in the Arctic. Nature, 411, 546-547. [JoC]
Sturm, M., T. Douglas, C. Racine and G.E. Liston, 2005: Changing snow and shrub conditions affect albedo with global implications. J. Geophys. Res., 110, G01004, doi:10.1029/2005JG000013. [JoC]
Syvitski, J.P.M., 2002: Sediment discharge variability in Arctic rivers: implications for a warmer future. Polar Res., 21, 323-330. [MoS]
Tannerfeldt, M., B. Elmhagen and A. Angerbjörn, 2002: Exclusion by interference competition? The relationship between red and arctic foxes. Oecologia, 132, 213-220. Clean
Tape, K., M. Sturm and C. Racine, 2006: The evidence for shrub expansion in Northern Alaska and the Pan-Arctic. Glob. Change Biol., 12, 686-702. [JoC]
Thomas, G. and P.R. Rowntree, 1992: The boreal forests and climate. Q. J. Roy. Meteor. Soc., 118, 469-497. Clean
Thompson, D.W.J. and S. Solomon, 2002: Interpretation of recent southern hemisphere climate change. Science, 296, 895-899. [PoC, JoC]
Thorpe, N., N. Hakongak, S. Eyegetok and the Kitikmeot Elders, 2001: Thunder on the Tundra: Inuit Qaujimajatuqangit of the Bathurst Caribou. Generation Printing, Vancouver, 220 pp. [NPR]
Trenberth, K.E., P.D. Jones, P.G. Ambenje, R. Bojariu, D.R. Easterling, A.M.G. Klein Tank, D.E. Parker, J.A. Renwick and Co-authors, 2007: Observations: surface and atmospheric climate change. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor and H.L. Miller, Eds., Cambridge University Press, Cambridge, 235-336. [NPR, PoC, 2007]
Truong, G., A.E. Palmé and F. Felber, 2007: Recent invasion of the mountain birch Betula pubescens ssp. tortuosa above the treeline due to climate change: genetic and ecological study in northern Sweden. J. Evol. Biol., 20, 369-380. [2007]
Tuck, G.N., T. Polacheck, J.P. Croxall and H. Weimerskirch, 2001: Modelling the impact of fishery by-catches on albatross populations. J. Appl. Ecol., 38, 1182-1196. [MoS]
Turchin, P. and G.O. Batzli, 2001: Availability of food and population dynamics of arvicoline rodents. Ecology, 82, 1521-1534. Clean
Turner, J., S.R. Colwell, G.J. Marshall, T.A. Lachlan-Cope, A.M. Carleton, P.D. Jones, V. Lagun, P.A. Reid and S. Iagovkina, 2005: Antarctic climate change during the last 50 years. Int. J. Climatol., 25, 279-294. [JoC]
Turner, J., J.E. Overland and J.E. Walsh, 2007: An Arctic and Antarctic perspective on recent climate change. Int. J. Climatol., 27, 277-293. [JoC, SRC, 2007]
Tutubalina, O.V. and W.G. Rees, 2001: Vegetation degradation in a permafrost region as seen from space: Noril’sk (1961–1999). Cold Reg. Sci. Technol., 32, 191-203. Clean
United States Environmental Protection Agency, 1997: Need for American Indian and Alaska Native Water Systems. U.S. Environmental Protection Agency, Washington, District of Columbia. [NPR]
U.S. Arctic Research Commission Permafrost Task Force, 2003: Permafrost, and impacts on civil infrastructure. 01-03 Arctic Research Commission, Arlington, Virginia, 62 pp. [NPR]
U.S. Geological Survey World Energy Assessment Team, 2000: Geological Survey World Petroleum Assessment 2000: Description and Results. U.S. Geological Survey Digital Data Series DDS-60, multidisc set, version 1.1, USGS. [NPR]
Usher, M.B., T.V. Callaghan, G. Gilchrist, B. Heal, G.P. Juday, H. Loeng, M.A.K. Muir and P. Prestrud, 2005: Principles of conserving the Arctic’s biodiversity. Arctic Climate Impact Assessment, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 540-591. [NPR, SRC]
van den Broeke, M. and N.P.M. van Lipzig, 2003: Response of wintertime Antarctic temperatures to the Antarctic Oscillation: results from a regional climate model. Antarctic Peninsula Climate Variability: Historical and Paleoenvironmental Perspectives, E. Domack, A. Leventer, A. Burnett, R. Bindschadler, P. Convey and M. Kirby, Eds., Antarctic Research Series 79, AGU, Washington, District of Columbia, 43-58. [NPR]
Van Oostdam, J., S. Donaldson, M. Feeley, N. Tremblay, D. Arnold, P. Ayotte, G. Bondy, L. Chan and Co-authors, 2003: Toxic substances in the Arctic and associated effects: human health. Canadian Arctic Contaminants Assessment Report II. Indian and Northern Affairs Canada, Ottawa, 82 pp. [NPR]
Van Wijk, M.T., K.E. Clemmensen, G.R. Shaver, M. Williams, T.V. Callaghan, F.S. Chapin III, J.H.C. Cornelissen, L. Gough and Co-authors, 2003: Long term ecosystem level experiments at Toolik Lake, Alaska and at Abisko, northern Sweden: generalisations and differences in ecosystem and plant type responses to global change. Glob. Change Biol., 10, 105-123. [JoC, SRC]
Vandenberghe, J., 2001: A typology of Pleistocene cold-based rivers. Quatern. Int., 79, 111-121. [SRC]
Vandenberghe, J., 2002: The relation between climate and river processes, landforms and deposits during the Quaternary. Quatern. Int., 91, 17-23. [SRC]
Vandenberghe, J. and M. Huisink, 2003: High-latitude fluvial morphology: the example of the Usa river, northern Russia. Paleohydrology: Understanding Global Change, K.J. Gregory and G. Benito, Eds., Wiley, Chichester, 49-58. [NPR, SRC]
Vaughan, D.G., 2006: Recent trends in melting conditions on the Antarctic Peninsula and their implications for ice-sheet mass balance. Arct. Antarct. Alp. Res., 38, 147-152. [SRC]
Vaughan, D.G., 2007: West Antarctic Ice Sheet collapse: the fall and rise of a paradigm. Climatic Change, in press. [NPR, JoC, SRC, 2007]
Vaughan, D.G., G.J. Marshall, W.M. Connolley, C.L. Parkinson, R. Mulvaney, D.A. Hodgson, J.C. King, C.J. Pudsey and J. Turner, 2003: Recent rapid regional climate warming on the Antarctic Peninsula. Climatic Change, 60, 243-274. [JoC, SRC]
Velicogna, I. and J. Wahr, 2006: Measurements of time-variable gravity show mass loss in Antarctica. Science, 311, 1754-1756. [JoC]
Vilhjálmsson, H., 1997: Climatic variations and some examples of their effects on the marine ecology of Icelandic and Greenland waters, in particular during the present century. Rit Fiskideildar, 15, 7-29. [SRC]
Vilhjálmsson, H., A. Håkon Hoel, S. Agnarsson, R. Arnason, J.E. Carscadden, A. Eide, D. Fluharty, G. Hønneland and Co-authors, 2005: Fisheries and aquaculture. Arctic Climate Impact Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 691-780. [NPR, SRC]
Vlassova, T.K., 2002: Human impacts on the tundra–taiga zone dynamics: the case of the Russian lesotundra. Ambio, 12, 30-36. Clean
Walker, M., H. Wahren, L. Ahlquist, J. Alatolo, S. Bret-Harte, M. Calef, T.V. Callaghan, A. Carroll and Co-authors, 2006: Plant community response to experimental warming across the tundra biome. P. Natl Acad. Sci. USA, 103, 1342-1346. [JoC, SRC]
Walsh, J.E., O. Anisimov, J.O.M. Hagen, T. Jakobsson, J. Oerlemans, T.D. Prowse, V. Romanovsky, N. Savelieva, M. Serreze, I. Shiklomanov and S. Solomon, 2005: Cryosphere and hydrology. Arctic Climate Impacts Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 183-242. [NPR, PoC, SRC]
Walsh, J.J., D.A. Dieterle and J. Lenes, 2001: A numerical analysis of carbon dynamics of the Southern Ocean phytoplankton community: the roles of light and grazing in effecting both sequestration of atmospheric CO2 and food availability to larval krill. Deep-Sea Res. Pt. I, 48, 1-48. Clean
Walter, K.M., S.A. Zimov, J.P. Chanton, D. Verbyla and F.S. Chapin, 2006: Methane bubbling from Siberian thaw lakes as a positive feedback to climate warming. Nature, 443, 71-75. [JoC]
Walters, C. and J.-J. Maguire, 1996: Lessons for stock assessment from the northern cod collapse. Rev. Fish Biol. Fisher, 6, 125-137. Clean
Walther, G.R., E. Post, P. Convey, A. Menzel, C. Parmesan, T.J.C. Beebee, J.M. Fromentin, O. Hoegh-Guldberg and F. Bairlein, 2002: Ecological responses to recent climate change. Nature, 416, 389-395. [JoC, ARC]
Warren, J., J. Berner and J. Curtis, 2005: Climate change and human health: infrastructure impacts to small remote communities in the North. International Journal of Circumpolar Health, 64, 498. Clean
Watt-Cloutier, S., 2004: Presentation to the Senate Committee on Commerce, Science and Transportation. September 15, 2004. Inuit Circumpolar Conference, Washington, District of Columbia. http://commerce.senate.gov/hearings/?testimony.cfm?id=1307&wit_id=3815 [NPR, MoS]
Weatherhead, B., A. Tanskanen, A. Stevermer, S.B. Andersen, A. Arola, J. Austin, G. Bernhard, H. Browman and Co-authors, 2005: Ozone and ultraviolet radiation. Arctic Climate Impact Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 151-182. [NPR]
Whinam, J. and G. Copson, 2006: Sphagnum moss: an indicator of climate change in the sub-Antarctic. Polar Rec., 42, 43-49. Clean
White, A., M.G.R. Cannel and A.D. Friend, 2000: The high-latitude terrestrial carbon sink: a model analysis. Glob. Change Biol., 6, 227-245. [JoC, MoS]
Wingham, D.J., A. Shepherd, A. Muir and G.J. Marshall, 2006: Mass balance of the Antarctic ice sheet. Philos. T. R. Soc. A., 364, 1627-1635. [JoC]
World Commission on Dams, 2000: Introduction to Global Change. Secretariat of the World Commission on Dams, Cape Town, 16 pp. [NPR]
Wrona, F.J., T.D. Prowse, J. Reist, R. Beamish, J.J. Gibson, J. Hobbie, E. Jeppesen, J. King and Co-authors, 2005: Freshwater ecosystems and fisheries. Arctic Climate Impact Assessment, ACIA, C. Symon, L. Arris and B. Heal, Eds., Cambridge University Press, Cambridge, 353-452. [NPR, SRC]
Wrona, F.J., T.D. Prowse, J.D. Reist, J.E. Hobbie, L. Levesque, R.W. Macdonald and W.F. Vincent, 2006a: Effects of ultraviolet radiation and contaminant-related stressors on Arctic freshwater systems. Ambio, 35, 388-401. [SRC]
Wrona, F.J., T.D. Prowse, J.D. Reist, J.E. Hobbie, L.M.J. Levesque and W. Vincent, 2006b: Climate change effects on Arctic aquatic biota, ecosystem structure and function. Ambio, 35, 359-369. [SRC]
Yang, D., B. Ye and A. Shiklomanov, 2004a: Discharge characteristics and changes over the Ob River watershed in Siberia. J. Hydrometeorol., 5, 595-610. [JoC]
Yang, D., B. Ye and D.L. Kane, 2004b: Streamflow changes over Siberian Yenisei River basin. J. Hydrol., 296, 59-80. Clean
Yang, D.Q., D.L. Kane, L.D. Hinzman, X. Zhang, T. Zhang and H. Ye, 2002: Siberian Lena River hydrologic regime and recent change. J. Geophys. Res., 107, 4694, doi:10.1029/2002JD002542. [JoC]
Yoccoz, N.G. and R.A. Ims, 1999: Demography of small mammals in cold regions: the importance of environmental variability. Ecol. Bull., 47, 137-144. Clean
Zhang, T.J., O.W. Frauenfeld, M.C. Serreze, A. Etringer, C. Oelke, J. McCreight, R.G. Barry, D. Gilichinsky, D.Q. Yang, H.C. Ye, F. Ling and S. Chudinova, 2005: Spatial and temporal variability in active layer thickness over the Russian Arctic drainage basin. J. Geophys. Res.–Atmos., 110, D16101, doi:10.1029/2004J D005642. [SRC]
Zhang, X. and J.E. Walsh, 2006: Toward a seasonally ice-covered Arctic Ocean: scenarios from the IPCC AR4 model simulations. J. Climate, 19, 1730-1747. [JoC, MoS, SRC]
Zöckler, C., 2005: Migratory bird species as indicators for the state of the environment. Biodiversity, 6, 7-13. Clean
Zwally, H.J., J.C. Comiso, C.L. Parkinson, D.J. Cavalieri and P. Gloersen, 2002: Variability of Antarctic sea ice 1979-1998. J. Geophys. Res.–Oceans, 107, 3041, doi:10.1029/2000JC000733. Clean
Zwally, H.J., M. Giovinetto, J. Li, H.G. Conejo, M.A. Beckley, A.C. Brenner, J.L. Saba and Y. Donghui, 2005: Mass changes of the Greenland and Antarctic ice sheets and shelves and contributions to sea-level rise: 1992–2002. J. Glaciol., 51, 509-527.Clean
| Tag | Explanation | Occurrences | Percentages |
|---|---|---|---|
| PoC |
Person of Concern Key individual involved in CRU emails as defined in this spreadsheet. |
9 | 2% |
| JoC |
Journal of Concern A Journal which has published articles by one or more PoCs (Person of Concern) |
105 | 29% |
| MoS |
Model or Simulation Reference appears to be a model or simulation, not observation or experiment. |
29 | 8% |
| NPR |
Non Peer Reviewed Reference has no Journal or no Volume or no Pages or it has Editors. |
114 | 31% |
| SRC |
Self Reference Concern Author of a chapter containing references to own work. |
74 | 20% |
| ARC |
Paper authored or co-authored by person who is also in list of Authors of another chapter. |
24 | 6% |
| 2007 |
Paper dated 2007, when IPCC policy stated cutoff was December 2005 |
11 | 3% |
| Clean |
The reference was probably peer reviewed. |
93 | 25% |
NOTE: In the Summary of Reviewer Comments,
Accepted includes only those responses that were unambiguously Accepted.
There are other categories of responses that we have not yet quantified
due to inconsistencies of usage.
| Reviewer | Total Comments | Accepted | IPCC Roles | Papers |
|---|---|---|---|---|
| Smith, SJ | 50 | 1 | 0 | 16 |
| Jefferies, RL | 48 | 3 | 1 | 2 |
| Atkinson, DE | 47 | 15 | 0 | 6 |
| Government of Norway | 47 | 4 | 0 | 0 |
| Government of Canada | 44 | 1 | 0 | 0 |
| McBean, GA | 38 | 3 | 1 | 3 |
| Palutikof, JP | 37 | 3 | 2 | 7 |
| Government of USA | 28 | 3 | 0 | 0 |
| Government of Poland | 25 | 1 | 0 | 0 |
| Cawkwell, Fiona | 23 | 0 | 0 | 0 |
| Parkinson, Claire | 18 | 0 | 0 | 0 |
| Streicker, John | 18 | 0 | 0 | 0 |
| Convey, P | 12 | 0 | 0 | 9 |
| Murray, T | 12 | 0 | 0 | 1 |
| van der Linden, Paul | 11 | 0 | 0 | 0 |
| Molau, U | 11 | 1 | 0 | 2 |
| Romanovsky, VE | 11 | 0 | 0 | 8 |
| Government of Netherlands | 8 | 0 | 0 | 0 |
| Meltofte, Hans | 8 | 1 | 0 | 0 |
| Hofgaard, A | 7 | 0 | 0 | 1 |
| Lange, Manfred | 7 | 0 | 0 | 0 |
| Government of Australia | 6 | 0 | 0 | 0 |
| Usher, Michael | 6 | 0 | 0 | 0 |
| Government of Finland | 6 | 0 | 0 | 0 |
| Government of Germany | 4 | 0 | 0 | 0 |
| Government of Sweden | 4 | 1 | 0 | 0 |
| Malcolm, David | 4 | 0 | 0 | 0 |
| Hare, W | 3 | 0 | 0 | 2 |
| Gajewski, K | 3 | 0 | 0 | 1 |
| Hindmarsh, Richard | 2 | 0 | 0 | 0 |
| Hansen, LJ | 1 | 0 | 0 | 4 |
| Government of Spain | 1 | 0 | 0 | 0 |
| Totals | 550 | 37 | 4 | 62 |
| Reviewer Type | Total Comments | Accepted | % Accepted |
|---|---|---|---|
| Govt | 173 | 10 | 5% |
| IPCC | 280 | 26 | 9% |
| Indep | 97 | 1 | 1% |
| Total | 550 | 37 | 6% |
Main AR4 Index | Working Group WG2 Index | Table of Contents | Authors | Executive Summary | Annotated Text | References | Reviewer Comments